
Genetyczne odkrycia 2010/2017 – Nowa Genealogia Słowian i innych ludów
Uwaga: TA Strona jest ciągle uaktualniana – jej stan się dynamicznie zmienia – Zapis bieżący na datę 6 sierpnia 2016 roku
Naprawdę warto odwiedzić strony histmagu i inne aktualne na 20 08 2016 roku .
Oto linki:
aktualna teoria na temat Genezy języków Indoeuropejskich 20 08 2016 – ze strony Tropie.tarnow.opoka.org.pl
DNA Forums: http://dna-forums.org/
Family tree DNA R1a: http://www.familytreedna.com/public/R1a/default.aspx?section=results
Histmag.org: http://histmag.org/forum/index.php/topic,10617.0.html
Kupując (TERAZ – TUTAJ!) Budujesz Wolne Media Wolnych Ludzi!

Przedstawiam najnowsze i jak się wydaje solidnie ugruntowane wiadomości na temat genetycznych odkryć dotyczących pochodzenia Słowian. Zacytowany tutaj rozdział 9 z opracowania dotyczy Wenedów i ich udziału w tworzeniu Słowiańszczyzny. Zainteresowanych głębiej tematyką genotypów i haplogrup odsyłam na stronę wyjściową tego materiału gdzie można znaleźć naprawdę gruntowne analizy poszczególnych zagadnień. Zamieszczone tutaj fragmenty pochodzą ze strony http://www.tropie.tarnow.opoka.org.pl/polacy.htm
Inna ważna strona na temat wpływu badań genetycznych na poglądy historyków i odczytanie przeszłych dziejów przez współczesną historię: http://histmag.org/forum/index.php?board=211.0

Mapa Kultur archeologicznych wytworzonych przez przedstawicieli poszczególnych haplogrup – z końca Epoki Brązu w Białym Lądzie (Europie). Mapa ta przedstawia jednocześnie przestrzeń zajmowaną przez poszczególne Ludy. Kultura Łużycka (na mapie Lusitian) oraz Thrace (tracka), Srubna i Kiukainen – to kultury Słowian lub Słowian i Dawian (Draków-Traków), oraz Słowian i Wenetów, albo Słowian i Indo-Irańczyków (Scytów-Skołotów i Sarmatów), a także Słowian i Bałtów. Na mapie widać ekspansję Słowiano-Dracką do Azji Mniejszej w okresie Upadku Troi. Na mapie poniżej opisano te same kultury (we wcześniejszym zasięgu) jak i wcześniejsze oraz równoległe kultury np. Trzciniecką przy pomocy haplogrup. Pokazano też ekspansję haplogrupy słowiano-istyjsko- skołocko-dawiańskiej (słowiano-bałtyjsko-scytyjsko-trackiej) na tereny Macedonii a następnie Grecji już w 1500 roku p.n.e., czyli w czasach przedtrojańskich. Czy to nie jest okres wojen Greków z Amazonkami i opisanej przez nich Amazonomachii? Czy nie są to Mazonki znad Morza Czarnego i Morza Mazońskiego (Azowskiego) i Mazonki z Mazowsza? Strzałka na mapie wychodzi spod północnych stoków Karpat – z Harii-Chrobacji Białej. Na poniższym opisie widać też że Kultura Łużycka powstała ze znacznym udziałem Słowian i Wenetów, a Słowianie mieli też duży wkład genetyczny w Kulturę Nordyczną, która później dała początek Germanom mieszając się z Celtami i Słowianami. Ludy Królestwa SIS weszły też w skład Kultury Minojskiej na Krecie, Iliryjskiej, Macedońskiej i Trackiej – R1a1a1 – to właśnie Ludy Królestwa SIS – Wielkiej Scytii, późniejszej Wielkiej Sarmacji i jeszcze późniejszej równie wielkiej Słowiańszczyzny. Przedstawieni tutaj Proto-Germanic People zajęli tereny Danii i i przyczółek na zachód od Półwyspu Duńskiego które wcześniej były zajęte przez Ludy R1a1a1 – czyli SIS. W skład celtyckiej kultury Tumulus także wchodziła grupa R1a – o czym warto pamiętać (warto- bo to są Ludzie z naszego późniejszego Weneca, a także współtworzyli tę kulturę Wenedowie).

-
„GENEalogia Y-DNA prawdę ci powie!” o początku i pokrewieństwie ludów na kuli ziemskiej (haplogrupy Y-DNA)
Oto pojawia się genetyka, a właściwie nie genetyka, lecz genealogia Y-DNA, ze swoimi fascynującymi możliwościami dla badaczy tych zagadnień! M.in. dostarcza ona danych, które umożliwiają umiejscowienie konkretnego człowieka na drzewie genealogicznym rodu, plemienia, wielkiego ludu lub nawet całej ludzkości.
To dlatego, że w męskim chromosomie Y przechowuje się i zasadniczo wiernie dziedziczy w trakcie przekazywania życia z ojca na syna materiał DNA wraz z jego historycznymi zmianami, zwanymi mutacjami.
Męski charakter genealogii Y-DNA jest istotny w ustalaniu pierwotnej grupy rodowej, plemiennej, czyli pierwotnego etnosu (gr. „etnos” to rój, trzoda, plemię). Bowiem w cywilizacji patriarchalnej ojcowski rodowód był istotnym czynnikiem genezy i rozwoju grup rodowych, plemiennych i etnicznych. Płciowy chromosom Y, choć nie służy przekazywaniu cech somatycznych (z wyjątkiem płciowych) ani tzw. rasowych, zawiera w sobie jakby bibliotekę rodowodu ojcowskiego wszystkich poprzednich pokoleń w linii prostej. W genealogii Y-DNA bierze się pod uwagę te mutacje, które dokonują się w niekodujących (niegenowych) odcinkach Y-DNA (ale to około 95% całej nici DNA!).
Mianowicie spośród około 57 milionów możliwych w chromosomie Y mutacji, niektóre, np. SNP (Single Nucleotide Polimophism; snip) dokonują się bardzo rzadko, bo w poszczególnych parach zasad (nukleotydów) przeciętnie raz na miliard lat, a więc praktycznie najwyżej raz w dziejach ludzkości i tylko u jednego mężczyzny, spontanicznie, jakby losowo, jako błąd w procesie kopiowania Y-DNA. Są one zasadniczo dokładnie przekazywane w trakcie zapłodnienia (mejozy) wszystkim męskim potomkom. Przy tym jednak, choć rzadko, może dojść do kolejnej mutacji, choć w innym miejscu (markerze), która zostaje przekazana potomstwu podobnie jak i poprzednie.
Mutacje SNP, przekazane męskim potomkom, są przez genetykę oznaczane odpowiednią literą, wskazującą na laboratorium, gdzie mutacje odkryto i cyfrą, oznaczającą kolejną liczbę odkrycia mutacji w tym laboratorium. Stąd np. takie symbole mutacji SNP: M417, L260, P45, CTS1211, DF93 itd.
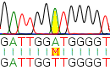 |
Komputerowy obraz mutacji typu SNP (single nucleotide polomorphism Symbole zasad/nukleotydów: A adenina, C cytozyna, G guanina, T tymina. Widzimy dwie sekwencje (dwa uklady) par zasad: wyżej, w próbce odniesienia jest sekwencja: GATTGGATGGGGT na dole, w próbce badanej sekwencja nieco inna: GATTGGTTGGGGT bowiem pojawiła się zasada T (tymina) tam, gdzie w próbce odniesienia (w linii na górze) jest A (adenina), czyli Adenina została zastąpiona Tyminą. Oznaczenie mutacji: A>T |
Od mutacji SNP zaczyna istnienie nowy ród (lub podród) genetyczny ojcowski (męski), a od mutacji mtDNA – genetyczny ród matczyny (żeński), zwany haplogrupą (h.g.) Y-DNA człowieka lub mtDNA (gr. 'haplous’ jednolity, pojedynczy). Haplogrupa więc to grupa ludzka, pochodząca od jednego mężczyzny lub kobiety, u których w trakcie poczęcia doszło do określonej mutacji zwanej SNP; mutacja ta występuje niezmiennie (gr. haplous) u wszystkich jego męskich potomków, nawet mimo pojawienia się u nich kolejnych mutacji (podobnie jak mutacje mtDNA u potomków żeńskich).
Haplogrupy oznaczamy umownie kolejnymi literami alfabetu łacińskiego, np. męskie – od A do T, oraz dodatkowo cyframi 1, 2, 3… oraz literami a, b, c… , jak np. R1a1a1 (jak aktualnie prasłowiańska i praindoirańska) albo R1b1a2 (jak podstawowa grupa genowa w Europie Zachodniej, zwana tez celtycką lub atlantycką).
Natomiast pierwszy mężczyzna, przodek wszystkich obecnie żyjących mężczyzn na świecie, tzw. Y-Adam, został oznaczony literą A. Potwierdziła się bowiem zapisana w Biblii tradycja o jednym człowieku u początku ludzkości, zwanym w genealogii genetycznej Y-chromosomalnym Adamem lub Y-Adamem. Pierwszego potwierdzenia dokonały badania Cruciani et al., maj 2011, który wykrył szereg mutacji SNP oznaczone później jako haplogrupa A1, dziedziczone głównie przez wszystkie haplogrupy pozaafrykańskie; oraz mutacje oznaczone jako bratnia haplogrupa A0, wykryte w Kamerunie i dziedziczone tylko przez tę grupę afrykańską. Z kolei w roku 2012 z inicjatywy grupy klientów komercyjnej firmy FTDNA (tzw. „citizen sciencists”) i na ich koszt dokonano kolejnej serii badań, które doprowadziły do wykrycia kolejnych osób z grupy A0, co umożliwiło zatwierdzenie wyników Crucianiego, a nadto daleko starszej haplogrupy, oznakowanej jako A00. Dziedziczna jest głównie przez nieliczną ludność żyjąca w południowo-zachodnim Kamerunie, wśród plemion Bangwa i Nkongho-Mbo. Paralelna do A00, bratnia grupa mutacji SNP, otrzymała oznaczenie A0T, gdyż jest dziedziczona przez wszystkie haplogrupy afrykańskie: A (oprócz A00) i B oraz pozaafrykańskie od C do litery T. Liczba SNP umożliwia datowanie tych haplogrup, a więc i ich wspólnego przodka, Y-Adama, na około 200.000 lat (w pracy opublikowanej przez Mendez et al. 2013 Y-Adama datowano nawet na około 338.000 lat!).
A oto laboratoryjna „metryka” jednej z tysięcy mutacji (SNP) Y-Adama, przekazanych obydwu liniom ludzkości – A00 i A0T – jako wynik zmiany zasady cytozyny w tyminę, czyli C->T. Odkryta jako pierwsza z tej grupy stała się symbolem Y-Adama (w 2012-2013 r. liczba znanych SNP Y-Adama wynosi już 1109):
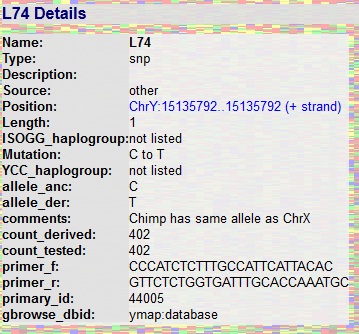
Powyższe badania umożliwiły opracowanie nowego diagramu filogenetycznego drzewa współczesnego człowieka.
| Drzewo genealogiczne współczesnej ludzkości. (otwórz w pdf) Stan wiedzy: 10 listopada 2012 i 2013 r. Porównaj: Nowe drzewo genealogiczne mtDNA |
Autor i kilku jego bliskich aktywnie uczestniczyli w inicjatywie, programowaniu i ponoszeniu kosztów testowania najstarszych, do niedawna nieznanych albo uważanych za odrębne od reszty ludzkości, rodów afrykańskich, celem ustalenia istnienia, czasu i miejsca chromosomalnego Adama, ojca całej dzisiaj żyjącej ludzkości. 10.11.2012 ogłoszono w USA wyniki badań klanu Perry’ego, rodem z Fontem w zachodnim Kamerunie. Jest to gałąź współczesnego człowieka, która najwcześniej wyłoniła się w rodzie Y-Adama. Dzięki temu przodka całej ludzkości, Y-Adama, datuje się obecnie albo na 338.000 (Hammer 2012; Mendez et al. 2013) lat, licząc jednak po 30 lat na jedno pokolenie (według dzisiejszego stanu). |
| Po koniecznej poprawce, na podstawie drzewa Scozzari et al. 2014, czas Y-Adama wynosi około 320.000 lat. Inne badania ukazują czas około 250.000 lat |

Aktualne drzewo genealogiczne genetycznych rodów ojcowskich
współczesnych ludzi. Wrzesień 2015 r.
– – –
wersja wcześniejsza:
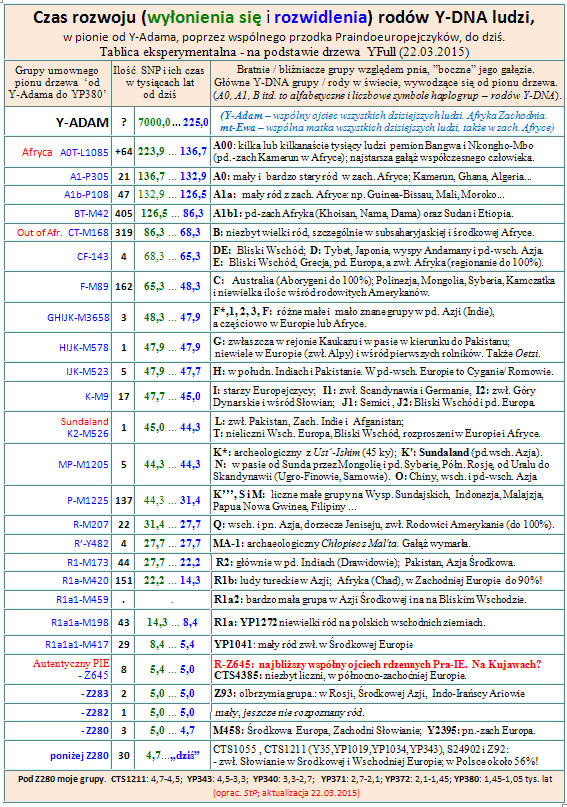
hg_czas.pdf
http://www.tropie.tarnow.opoka.org.pl/images/hg_time.jpg
hg_time.pdf
Zobacz procentowe rozmieszczenie poszczególnych haplogrup na mapie świata.
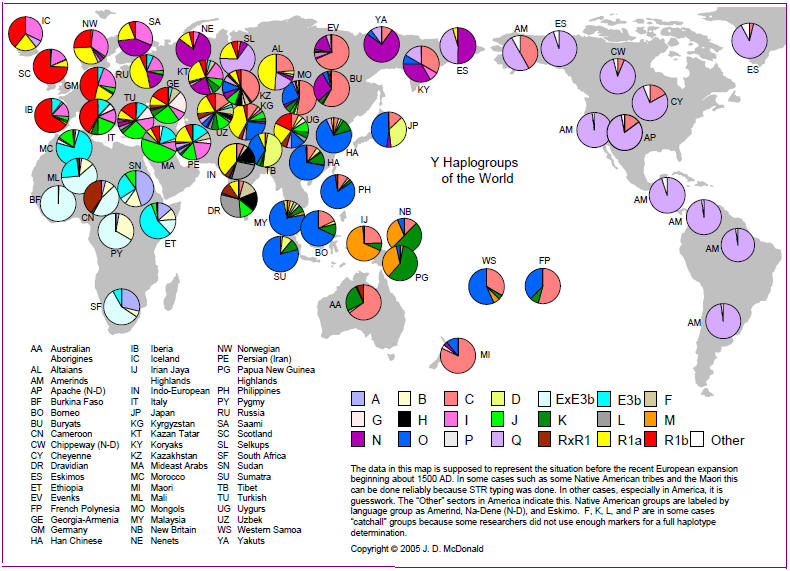
http://www.scs.illinois.edu/~mcdonald/WorldHaplogroupsMaps.pdf
oraz: mapy rozmieszczenia w świecie poszczególnych haplogrup
Por. datowanie węzłów międzygrupowych (Marco Heinla, 2011): http://beforepresent.dyndns.info/timevalues.html
oraz sporządzone przez Dienekesa Pontikosa i opublikowane na jego blogu:
http://dienekes.blogspot.com/2012/08/dates-of-major-clades-of-y-chromosome.html
………………………………………..
Potwierdziła się także tradycja o wspólnej matce wszystkich żyjących na świecie kobiet. Bowiem dzięki mechanizmowi wiernego przekazywania potomstwu tylko przez matkę genu mitochoindrialnego, mt-DNA, regulującego powstanie i działanie mitochondriów komórkowych, służących „energetyce organizmu” oraz dzięki mutacjom, które w trakcie kopiowania w tym genie zachodzą, można dojść do pierwszej matki w linii prostej, czyli tzw. mitochondrialnej Ewy. W genealogii genetycznej datuje się ją obecnie na około 177 tysięcy lat temu (Behar et al. 2012) i też w środkowo-wschodniej Afryce.
Zespół mutacji mitochondrialnej Ewy oznaczono symbolem
RSRS
– skrót od Reconstructed Sapiens Reference Sequence.
W ten sposób, korzystając ze znajomości drzewa genealogicznego mutacji w Y-DNA i w mtDNA, można genealogię swojego rodu, znaną np. z dokumentów i tradycji rodzinnej, uzupełnić genealogią genetyczną – aż do pierwszych praojców: chromosomalnego „Adama” i mitochondrialnej „Ewy”.
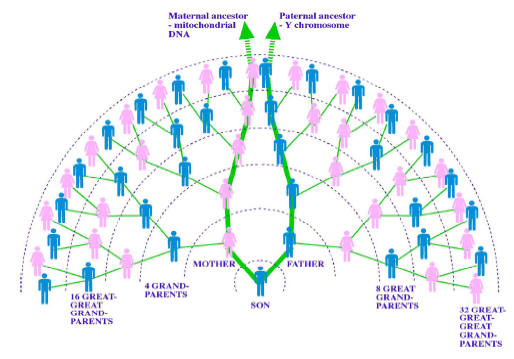 |
Ilustracja dziedziczenia
genów mitochondrialnych (mtDNA) tylko w prostej linii matczynej |
http://www.nottingham.ac.uk/-sczsteve/Gothenburg_13Oct2011.pdf
Zob. przykład takiej genealogii ojcowskiej Y-DNA i matczynej mtDNA pewnej rodziny.
Ze względu na to, że w naszej cywilizacji patriarchalnej (ojcowskiej, męskiej) pierwotne grupy etniczne mają genezę patriarchalną, co potwierdza archeogenetyka, interesuje nas szczególnie ustalenie pokrewieństwa i pochodzenia wspólnot plemiennych i etnicznych przez śledzenie dziedzictwa genetycznego w męskim chromosomie Y-DNA. Zajmuje się tym genealogia Y-NA, zwana też, choć trochę niewłaściwie, genogenealogią. Genealogia Y-DNA bywa w antropologii niekiedy mylona z pokrewną nauką, zwaną genetyką populacyjną, której zadaniem jest śledzić genetyczne podobieństwa między dzisiejszymi społecznościami, z całą ich biologiczno-genową złożonością.
Czas rozwoju (wyłonienia i rozwidlenia) głównych haplogrup: hg_czas.pdf
English: hg_time.pdf
Odkrycia i datowanie „Człowieka z Ust’-Ishim” u uścia’ (sic!) rzeki Iszym
do Irtyszu w zachodniej Syberii, a także haplogrupy K2* (Fu et al., 2014)
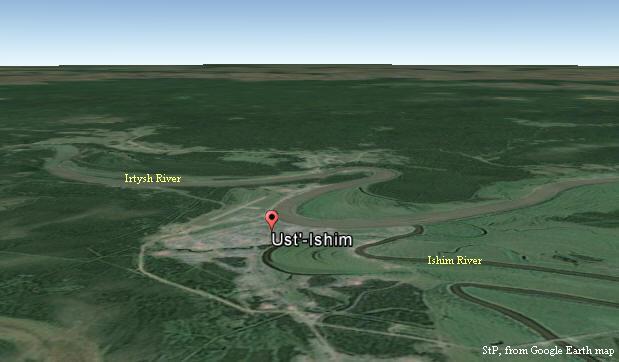
Tam, w Ust’-Ishim nad rzeką Irtysz, 45.000 lat temu żył „Czlowiek z Ust’-Ishim”, potomek haplogrupy K2*, żyjącej około 50.000 lat temu, która wcześniej migrowała w kierunku Sundalandu, gdzie wnet pojawił się inny jej potomek, haplogrupa MP, praprzodek większości Europejczyków (zob. tabela Czasu rozwoju haplogrup Y-DNA).
Tempo mutacji (SNP) w Y-DNA 0,76×10^-9
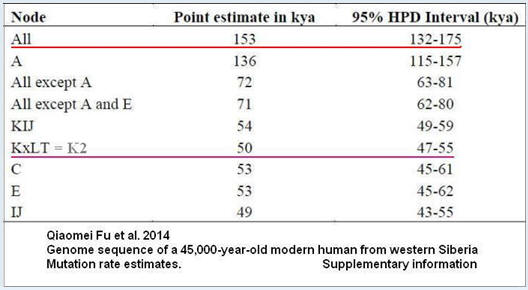
Uwaga.
Zaniżone datowanie wszystkich (All) haplogrup człowieka współczesnego (153 tysiące lat zamiast około 275 lub przynajmniej 200 tysięcy, jak do niedawna datowano) jest wynikiem braku pełnego testowania ludzi haplogrupy A00.
- 2.
Genealogia Y-DNA POWIE CI też o bliższym pokrewieństwie
rodów, rodzin i osób
(Haplotypy Y-DNA)
Oprócz omówionych wyżej mutacji SNP, w męskim Y-DNA dokonują się jeszcze inne, mniej czytelne w genealogii, ale daleko częstsze zmiany. Polegają one na zmiennej ilości powtórzeń odpowiedniego tandemu (sekwencji) zasad, np. ATGC, w pozagenowych mikrosatelitarnych markerach Y-DNA, nazywanych STR (Short Tandem Repeats, czyli powtarzenia krótkiego tandemu, czyli sekwencji, zasad A-T-G-C). Ulegające zmianom miejsca, w których dokonują się owe powtórzenia, nazywane są „loci” lub „markery” i oznaczane są skrótem Dys# i numerem tego miejsca; zaś zmieniona liczba powtórzeń owych tandemów stanowi liczbę alleli w danym markerze (miejscu), np. DYS393=13. Zestaw liczb owych powtórzeń (alleli) u jednego mężczyzny stanowi jego haplotyp (ht) STR. Haplotyp, czyli zestaw ilości mutacji w poszczególnych miejscach (markerach) Y-STR jednego mężczyzny, umożliwia porównanie z haplotypem innego mężczyzny z tej samej haplogrupy, przeliczenie dzielących ich mutacji czyli GD (genetyczny dystans), ustalenie na ich podstawie wspólnego najbliższego przodka (MRCA) i rozpoznawanie bliskości ich pokrewieństwa. Bywa to wykorzystywane np. w sądownictwie (dla ustalenia lub wykluczenia ojcostwa) albo w budowie drzewa genealogicznego jakiegoś wielkiego rodu.
Spotykane liczby alleli w poszczególnych 67 markerach (loci), w standardowej kolejności firmy FTDNA i w najczęstszych haplogrupach w Europie przedstawione są na na specjalnej tablicy, a zwłaszcza na naszej stronie: Dyskusja o mutacjach.
Od 5 kwietnia 2011 r. jako standard uważa się testowanie nie 67, a 111 markerów w firmie FamilyTree DNA.
Zestaw podobnych haplotypów STR może być niekiedy użyty bez bardziej kosztownych badań, choć z daleko mniejszą pewnością, do ustalenia haplogrupy SNP, w ramach której owe haplotypy powstały. Jednak trzeba wiedzieć, że podobne haplotypy mogą pojawiać się równolegle w rozmaitych haplogrupach.
Oto przykład 25-markerowego haplotypu modalnego, czyli najczęstszego, słowiańskiej haplogrupy R1a1a w krajach Europy Środkowej i Wschodniej: zwłaszcza: Polska, Ukraina, Rosja, Białoruś, Litwa, Niemcy, Czechy, Słowacja i Węgry (według Wikipedii)
| DYS | 393 | 390 | 19 | 391 | 385A | 385B | 426 | 388 | 439 | 389I | 392 | 389II | 458 | 459A | 459B | 455 | 454 | 447 | 437 | 448 | 449 | 464A | 464B | 464C | 464D |
| Allele | 13 | 25 | 16 | 10 | 11 | 14 | 12 | 12 | 11 | 13 | 11 | 30 | 16 | 9 | 10 | 11 | 11 | 23 | 14 | 20 | 32 | 12 | 15 | 15 | 16 |
Liczba DYS w programach FTDNA oznacza numerowane miejsce (marker) w STR męskiego Y-DNA.
Allele wyrażają ilość powtórzeń określonej sekwencji (tandemu, następstwa) zasad A, C, G, T w danym miejscu Y-DNA.
Np. na podstawie Dys 385a=11, 439=11, 447=23 rozpoznaje się powyższego haplotypu haplogrupę R1a1a1g, typ N (kraje słowiańskie).
Powstanie w drodze mutacji (np. przestawienia kolejności zasad A,C,G,T) nowych haplotypów w ramach jednej haplogrupy wymaga czasu. Stąd ilość tych mutacji w Y-STR bywa wykorzystywana w ramach genealogii Y-DNA do obliczania czasu od wspólnego przodka, w skrócie TMRCA (Time to the Most Recent Common Ancestor). Metodami statystycznymi ustalono, że:
1) w przypadku przeliczenia alleli w 12-markerowym haplotypie należy się spodziewać 0,022 mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na 1136 lat.
2) w przypadku przeliczenia alleli w 25-markerowym haplotypie należy się spodziewać 0.046 mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na 543 lata.
3) w przypadku przeliczenia alleli w 37-markerowym haplotypie należy spodziewać się 0.09 mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na około 278 lat.
4) w przypadku przeliczenia alleli w 67-markerowym haplotypie należy spodziewać się 0,12 mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na około 208 lat (stawki Klosowa-Rożanskiego w: Wiestnik 3/12, 2010 r.).
| UWAGA! Po sprawdzeniu wielu zagranicznych publikacji naukowych i śledząc naukowe fora poświęcone genealogii Y-DNA, zwłaszcza RootsWeb, GeneDiversitas, MolGen lub Rodstvo, dochodzę do przekonania o dość znacznej wiarygodności rosyjskiego środowiska naukowego, któremu przewodzi rosyjsko-amerykański naukowiec, prof. Harwardu Anatol KLOSOW (ang. Anatole Klyosov, ros. Анатолий A. Клёсов, tu w skrócie: A.K.; Moskwa-Boston-Massachussetts), biochemik, specjalista od kinetyki chemicznej, autor uniwersyteckiego podręcznika w tej dziedzinie oraz bardzo licznych i całościowych prac z dziedziny genealogii Y-DNA. Obecnie z powodzeniem udoskonala i propaguje on naukowe zasady liczenia mutacji STR, budowy genealogicznego drzewa Y-DNA i obliczania czasu do wspólnego przodka populacji danej haplogrupy (TMRCA). Ostatnio w analizie haplotypów i ostrożnym datowaniu populacji wyróżnia się też naukowiec-biochemik japoński ukraińskiego pochodzenia, Igor Rożanski, cieszący się środkowoeuropejską haplogrupą R1a1a1g-M458. Jego dziełem jest zestaw cennych analiz i map drzewa genealogicznego R1a1a1, z których korzysta się na tej stronie. |
| Wykładniczy czas mutacji widocznych (obserwowalnych)
Dotąd bardzo trudny problem rozpoznawania ilości mutacji powtórnych / zwrotnych, pojawiających się w markerach szybciej mutujących, zupełnie niewidocznych i niedających się odczytać, który zmuszał autorów do wykonywania rozmaitej akrobatyki matematycznej, stara się rozwiązać najnowsza tabela Pełnego Czasu Mutacji ( PCM / FTM, zob.), utworzona na naszych stronach i opublikowana na niektórych międzynarodowych forach naukowej genealogii genetycznej. |
Obecnie każdy człowiek, przekazując wymaz (z jamy ustnej) ze swoim DNA, może wziąć udział, z niewielkim wydatkiem pieniężnym, w programie tych badań (program FamilyTreeDNA). Zob. Polish project, który administruje Amerykanin polskiego pochodzenia, L. Mayka. Krótki poradnik zob. niżej, w rozdziale 16.
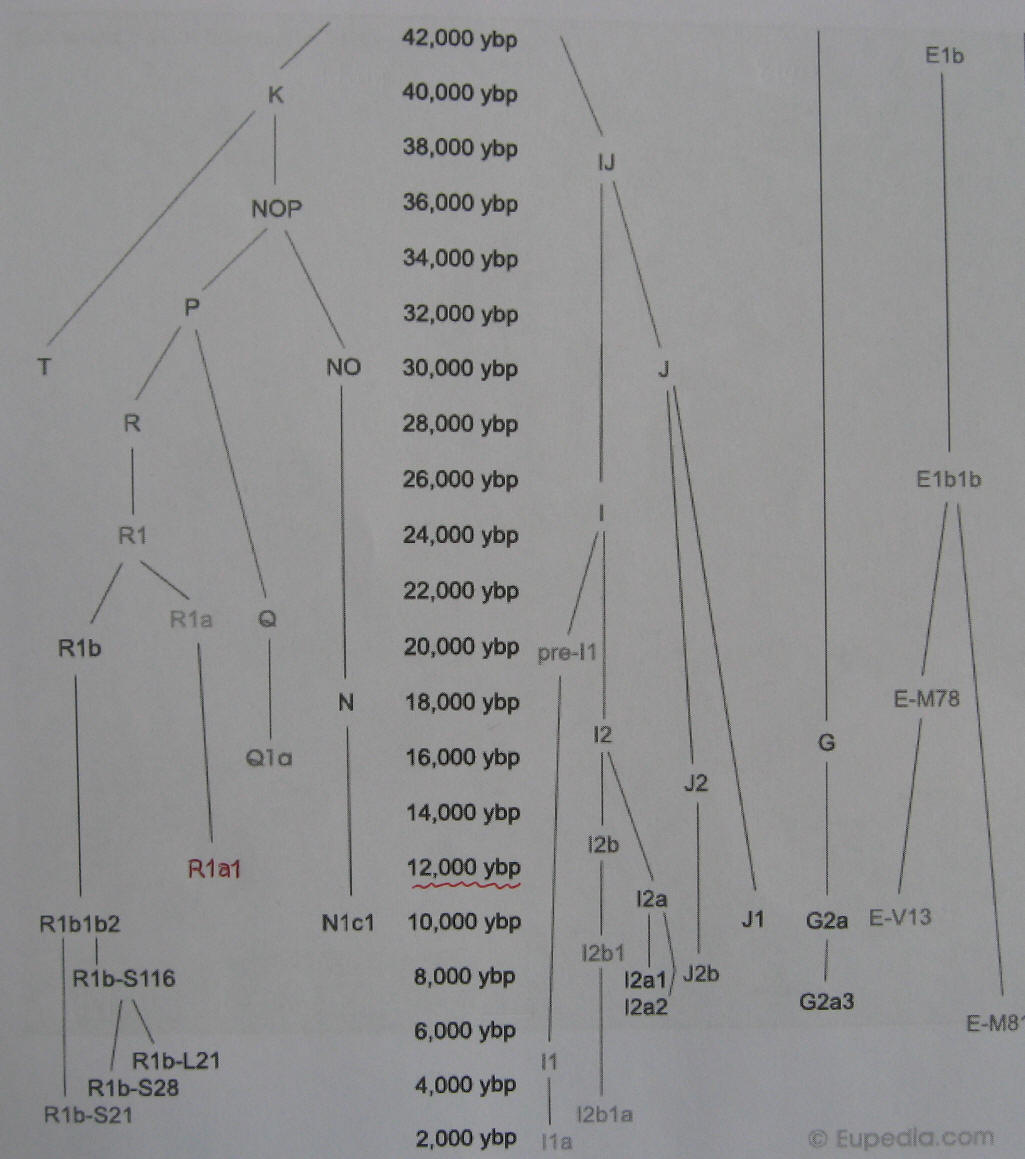 |
Dolna część drzewa genealogicznego ludów Europy, tylko haplogrupy męskie Y-DNA. Od lewej – haplogrupy dzisiejszych ludów: R1b, R1a1, Q1a, N1c1, I1a, I2a2, J2b, J1, G2a3, E-V, E-M. O nich mowa niżej. W centrum – czas powstania (w latach) odpowiednich mutacji Y-SNP czyli początku poszczególnych haplogrup. (Eupedia-tree2) |
3.
jak powstały ludy pozaafrykańskie?
Mapa ogólna rozprzestrzeniania się współczesnego człowieka z Afryki do Eurazji i dalej.
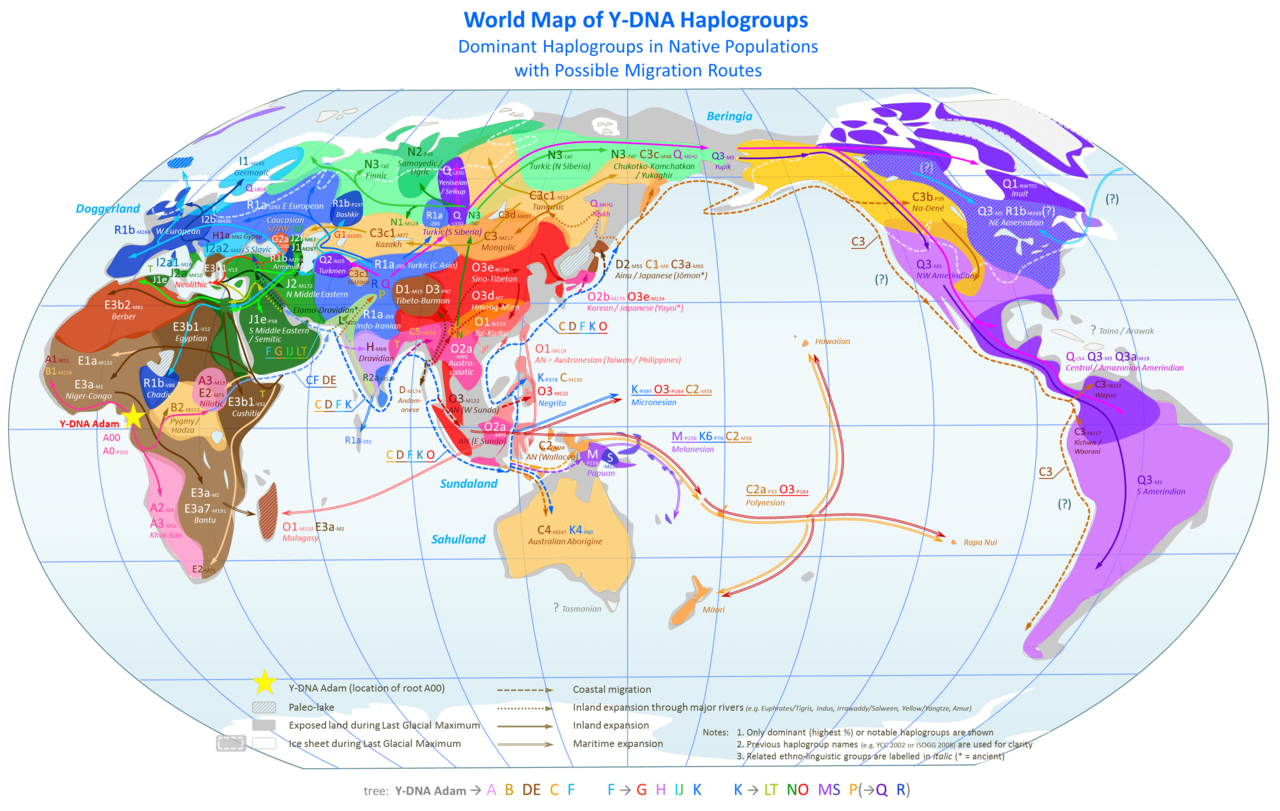
Migracje i rozmieszczenie haplogrup Y-DNA w świecie (kliknij, powiększ!)
Zob. miejsce haplogrupy A00 i A0 w Kamerunie; tam początek współczesnego człowieka.
źródło: https://en.wikipedia.org/wiki/File:World_Map_of_Y-DNA_Haplogroups.png
Zob. pochodzenie rdzennych Amerykanów,Ri Q: http://www.sciencemag.org/content/342/6157/409.full
(Zobacz też: Pochodzenie ludności w rejonie Dunajca)
https://dnaexplained.files.wordpress.com/2014/10/hammer-2014-30.jpg?w=584&h=380
Pierwsze słynne „Out of Africa” współczesnego człowieka na Półwysep Arabski mogło dokonać się wnet po 100.000 lat temu, o czym mają świadczyć niektóre świadectwa archeologiczne. W tym okresie, zapewne w Afryce, powstaje haplogrupa BT*, której liczne SNP są dziedziczone przez wszystkie pozaafrykańskie haplogrupy. Na czas około 70.000 lat temu datuje się wielką katastrofę przyrodniczą, związaną z wybuchem wulkanu Toba w Indonezji. Musiała ona dotknąć i najstarsze populacje ludzkie, migrujące w kierunku Azji. Widoczne jest to także na haplogrupie BT, której liczne mutacje w jednej linii ojcowskiej świadczą o przeżytym wąskim gardle (bottleneck) demografii. Przeżył tylko jeden przodek, oznaczony dziś jako haplogrupa BT, który otrzymał szansę być ojcem prawie całej pozaafrykańskiej ludności świata.
Zapewne poza Afryką, około 70.000 lat temu, już poza Afryką haplogrupa CT rozgałęziła się na DE i CF. Hg E – po wydzieleniu siĘ wnet powróciła do Afryki, gdzie się bardzo upowszechniła i stanowi większość ludów tego kontynentu. Hg D – migrowała daleko do Azji środkowej i wschodniej (Japonia). Potomkowie z hg C żyją w krajach Azji zachodniej, południowej i południowo-wschodniej, nadto jako aborygeni w Australii oraz pierwotna ludność na wyspach Polinezji i nielicznie w Ameryce.
Natomiast potomstwo naszego przodka z haplogrupą F dało początek prawie 90% pozaafrykańskiej ludności świata. W gronie potomków haplogrupy F (naszego przodka, zob.) w Azji Mniejszej wyłoniła się najpierw współna haplogrupa GHIJK, a wnet potem żyjąca dziś głównie na Zakaukaziu i nielicznie w Europie haplogrupa G (wyróżniająca dziś głównie Ormian). Haplogrupa G na początku VI tysiąclecia przed Chr. przyniosła do Europy rolnictwo. Natomiast z synowskiej haplogrupy HIJK wyłoniła się haplogrupa H. To żyjący dziś w Indiach, a potem ich część także w Europie jako Cyganie/Romowie.
Od około 55 tysięcy lat temu Azji Mniejszej z ojcowskiej grupy IJK rozwija się populacja z haplogrupą IJ, a z niej potem wyłoni się w Europie ludność staroeuropejska, „przedindoeuropejska”, z haplogrupą I, którą by trzeba utożsamić prawdopodobnie ze znanymi w Europie populacjami przeważnie pod nazwą Wenetowie/Wenedowie. W południowej Skandynawii dali oni początek etnosowi pragermańskiemu. Natomiast w Azji Mniejszej powstała haplogrupa J, głównie semicka.
W czerwcu roku 2014 T. Karafet et al. ogłosili wyniki badań ważnych dla ustalenia, dotąd raczej enigmatycznie nakreślanego, drzewa filogenetycznego w pionie haplogrup A-R, w jego młodszych gałęziach: Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia, (Link i dostęp w: http://dienekes.blogspot.com/2014/06/refined-structure-in-haplogroup-k-m526.html ).
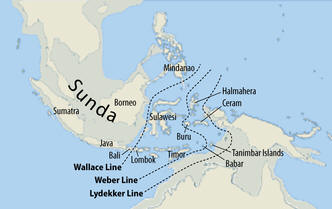
Mianowicie, w rodzie naszego praprzodka z haplogrupą K-M9, po wydzieleniu się gdzieś w Azji Poludniowej podgrupy K1-P326, w której powstały gałęzie L (żyjące dziś zwł. w Pakistanie, Indiach i Sri Lance) i T (żyjąca dziś na Bliskim Wschodzie, pd.-zach. Afryce i w Europie), w drodze ku Azji Południowo-Wschodniej wyłoniła się najpierw mutacja K2-M526 (bliski przodek „człowieka a UST’-Ishim”, datowanego radiowęglowo na 45 tysięcy lat), a już zapewne na terenie Sundalandu, (dziś Wyspy Sundajskie w Indonezji) wówczas w całości będącego na powierzchni, kolejna podgrupa z mutacją P331, oznaczana teraz jako haplogrupa K2b, oznaczana także jako MP.
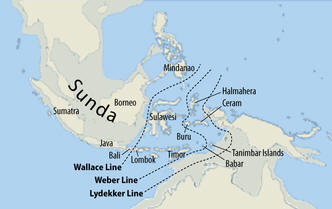
Sundaland i jego domniemane granice południowe,
stosownie do zmiennego poziomu mórz ( http://en.wikipedia.org/wiki/Sundaland )
Haplogrupa MP uległa tam szybkiemu rozwojowi demograficznemu. Zapewne pod koniec okresu wielkiego zlodowacenia (LGM) około 18-15 tys. lat temu i podniesienia się poziomu mórz zaszła konieczność ucieczki z Sundalandu i emigracji euroazjatyckiej, w której brały udział powstające haplogrupy Q i R.
Oto jak M. Hammer przedstawia migrację od Sundalandu ku Europie:
https://dnaexplained.files.wordpress.com/2014/10/hammer-2014-30.jpg?w=584&h=380
Niektórzy potomkowie haplogrupy Q poprzez północno-wschodnią Syberię i Alaskę około 15.000 lat temu zawędrowali na kontynent Amerykański, dając wraz z hg C początek autochtonnym Indianom w Ameryce Północnej i Południowej.
Natomiast haplogrupa R-M207 powstała około 34.000 lat temu (zob. tabela) i rozproszyła się gdzieś m.in. w rejonie Ałtaju. O tym świadczy jej potomek, tzw. „Chłopiec z Malta (Mal’ta)”, którego archeologiczne szczątki, datowane na 24.000 lat temu, znaleziono we wsi Mal’ta, w pobliżu Jeziora Bajkał. W tym rejonie mogło też dojść do wydzielenia się i migracji podgrup R1 i R2. Ta druga zatrzymała się głównie na Półwyspie Indyjskim. Ta pierwsza, z mutacją M173 i swoimi podgrupami R1a i R1b, rozproszyła się głównie w Eurazji Środkowej i Zachodniej, głównie w Europie.
 |
MALTA. Na Syberii, pod południową granicą Federacji Rosyjskiej, nad wypływającą jako jedyną z jeziora Bajkał rzeką Biała (wpływa do rzeki Angara, Jeniseju do Morza Karskiego i Arktyki, poniżej miasta Irkuck znajduje się wioska Malta. Odkryto tam paleolityczny grób z roku około 24.000 lat temu z czaszką chłopca i jego genetyczną haplogrupą R (bez znanych nam młodszych mutacji w haplogrupzch R1, R2 i R1a/R1b): http://antropogenez.ru/article/743/Dziś w okolicy Malta żyje syberyjskie niewielkie plemię Keti, http://www.narodsevera.ru/north/narod/keti które dziedziczy zapewne cechy tamtejszej ludności sprzed 24 tysięcy lat, a może także niektóre jej obyczaje.http://dienekes.blogspot.com/2013/11/ancient-dna-from-upper-paleolithic-lake.html |
Populacje haplogrupy R1 z jej podrupami R1a i R1b, które zachowały pierwotny „ojcowski” język, są twórcami i użytkownikami języków tzw. indoeuropejskich, nazwanych tak ze względu na geograficzną ich rozległość od Subkontynentu Indyjskiego na południu Azji po zachodnie krańce kontynentu Europejskigo.
Uwaga. Powyższe dane porównaj: Chiaroni et al. 2009; tam też najnowsza mapa powstania poszczególnych haplogrup, ich migracji oraz aktualnego rozmieszczenia ich populacji w świecie (mapa).
Główne haplogrupy, R1a (kolor pomarańczowy) i R1b (kolor czerwony) i inne w Europie
i na terenach przyległych na południu i południowym wschodzie; według wikipedii, rok 2013.
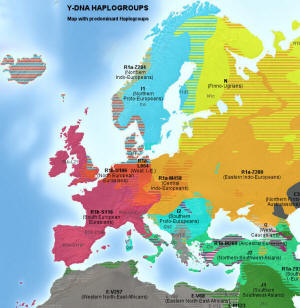 Główne haplogrupy Y-DNA w Europie Główne haplogrupy Y-DNA w Europie(kliknij, aby powiększyć) |
Kolor żółty: haplogrupa N (N1b i N1c) – grupy ugrofińskie
Pomarańcz.: haplogrupa R1a, grupa rdzennie indoeuropejska Niebieski: haplogrupa I (staroeuropejska) Czerwony: haplogrupa R1b zachodnioeuropejska, Zielony: haplogrupa J, bliskowschodnia, nieindoeuropejska Szary: haplogrupa E, afrykańska; E-V13 bałkańska |
5.
Haplogrupa/RÓD R1a i jego główne podgrupy.
Haplogrupy Słowian i ich najbliższych krewnych, indo-irańskich Ariów.
Z genezą Słowian jako etnosu (gr. „etnos” – plemię, ród) sprawa jest niejasna, podobnie jak z innymi starożytnymi ludami. Na podstawie opisów, jakie zostawili nam starożytni pisarze, ale dopiero począwszy od Herodota (V w. przed Chr.), Słowianie odznaczali się trzema głównymi cechami: „Po pierwsze: lokalizacja gdzieś w dorzeczu Dniepru, Dniestru, po Karpaty lub ewentualnie ujście Dunaju. Po drugie: usytuowanie na terenach o dobrze rozwiniętej sieci wód śródlądowych, gęstych lasach, bagnach, na obszarach wybitnie nizinnych. Po trzecie: muszą być w sensowny sposób związane z młodszymi kulturami archeologicznymi z okresu wczesnego średniowiecza, dla których stanowiły podstawę uformowania sie i dalszego rozwoju” (A. Błażejewski). Na tej podstawie Słowianom można by przypisać archeologiczne kultury wysocką i wołyńską nad górnym Bugiem i Dniestrem (może to dzieła Herodotowych Neurów i Budynów z około V w. przed Chr.), a już na pewno zarubiniecką i czerniachowską (od III w. przed Chr.) oraz kijowską (od III w. po Chr.), na Ukrainie. Najstarszą słowiańską wersją ich nazwy zanotował Prokopius z Cezarei: Spori, która skojarzyła sie mu ze słowem sporadoi 'sporadyczni, rozproszeni’. Ale bliższa analiza językoznawcza już wykazała, że słowiańskie słowo spori oznaczało „sporzy, liczni”.
(Niestety, złą przysługę prawdzie naukowej wyrządzili ostatnio skrajni allochtoniści, zwani kossinowcami-dnieprowcami, którzy – na podstawie zacieśnionej definicji archeologicznej kultury słowiańskiej – początek Słowian na ziemiach datują jedynie na VI w., ich praojczyznę lokują jedynie nad środkowym i górnym Dnieprem, a ich genezę ustalają tam na czas niewiele wcześniejszy. Za nimi poszli niektórzy polscy genetycy z kręgu medycyny sądowej, np. K. Rębała i zesp. 2007, którzy badając pochodzenie dzisiejszych Polaków, nie rozróżniają ich chromosomowych haplogrup SNP, a biorąc pod uwagę tylko haplotypy STR, nieraz bardzo podobne mimo odległych pochodzeniem haplogrup SNP, oceniają ich podobieństwo czy różnorodność względem haplotypów innych populacji i na tej podstawie orzekają o dacie ich genezy w… V czy VI w. i miejscu ich oddzielenia się od innych ludów – nad Dnieprem! Podobna metoda, oparta na mieszaniu haplotypów rozmaitych haplogrup i korzystaniu także z materiału autosomalnego, zawiodła już wcześniej R. Płoskiego i zesp. 2002 do stwierdzenia, wbrew naukowej genetyce, o wielkiej odrębności Polaków od innych ludów słowiańskich; że pod „względem genetycznym jesteśmy prawie tak samo oddaleni od Moskwy, jak i od Berlina”).
W Rodzie R1a, powstałym zapewne w zji Południowo-Wschodniej, na terenie dawnego Sundalandu (zob. też niżej, tabela mutacji), w trakcie migracji ku Europie lub już w Europie (Kujawy?) powstała kolejna mutacja SNP, oznaczona symbolem M417 i określana obecnie jako haplogrupa R1a1a1, uważana jest za ojcowską dla indoeuropejskich ludów; bowiem pod nią doszło do podziału tej rodziny genetyczne na dwie wielkie grupy. Jedna, z mutacją Z93, dała początek indo-irańskim i azjatyckim Ariom i ich językowej odrębności. Druga zaś, z mutacja Z283 dała początek wielu ludom europejskim, a zwłaszcza Bałto-Słowianom i ich językom. Niektórzy naukowcy w populacjach Z283 widzą genezę wszystkich indoeuropejskich języków w Europie, także tych w europejskich haplogrupach R1b.

Rzecz w tym jednak, że rzetelna genetyczna genealogia Y-DNA i językoznawstwo historyczne nie pozwalają na takie, jak wyżej, czasowe, terytorialne i kulturowe zacieśnienie definicji Słowian starożytnych. Poucza o tym analiza czasu ich powstania oraz geograficznego rozprzestrzenienia się populacji Słowian, wyróżniającej się własnym językiem i genową haplogrupą R1a1a1 (znana do niedawna jako R1a1a). W listopadzie 2009 z tej haplogrupy wydzielono ważną podgrupę, oznaczaną dziś jako R1a1a1g.
Otóż w populacji hg R1 (M173), oprócz wyżej omówionej „italo-celtyckiej” M343 (haplogrupa R1b), dokonała się też mutacja M420, dająca początek ważnej indo-europejskiej haplogrupie R1a. Jej potomków, bez kolejnych mutacji, czyli paragrupę R1a* badania Underhilla, obejmujące 7224 osób w Eurazji, zidentyfikowały w kilku izolowanych regionach Bliskiego Wschodu i Kaukazu, łącznie siedem osób w Iranie, Omanie, Emiratach Arabskich i Turcji. W rejonie więc Bliskiego Wschodu należy upatrywać kolebki wielkiej, liczącej dziś ponad 270 milionów populacji R1a1a i jej podgrup w Eurazji.
Kolejna mutacja, M448 (SRY1532.2) w haplogrupie R1a-M420, określa haplogrupę/podgrupę R1a1. Ta haplogrupa nie zostawiła po sobie liczniejszych populacji. Badania M. Requeiro i zesp. 2006, wskazują na Iran jako miejsce pochodzenia hg. R1a1. Tam M448 zidentyfikowano ogólnie w 3% badanych, a w niektórych plemionach północnego Iranu – nawet 15% (co jednak wymaga weryfikacji). Potwierdzałoby to wysunięte przypuszczenie o bliskowschodniej kolebce R1a i R1a1, czy dokładniej – Płaskowyż Irański, jak wskazuje W. Ryżkow na podstawie swoich badań paleokraniometrii.
Natomiast po kolejnych mutacjach (M17/M198) – powstała euro-azjatycka haplogrupa R1a1a, a pod nią – niedawno rozpoznana wielka haplogrupa M417, określana jako R1a1a1. Potem w jej części europejskiej doszło do kilku mutacji L664 (nieliczni jej nosiciele zidentyfikowani w Europie Pólnocno-Zachodniej), M458, która utworzyła skoncentrowaną wśród Słowian środkowoeuropejskich haplogrupę R1a1a1g oraz mutacji L260. Tę ostatnią zdefiniowano jako zachodniosłowiańską podgrupę R1a1a1g2, obecną szczególnie w Polsce, Czechach i Słowacji i określaną niekiedy jako polską.
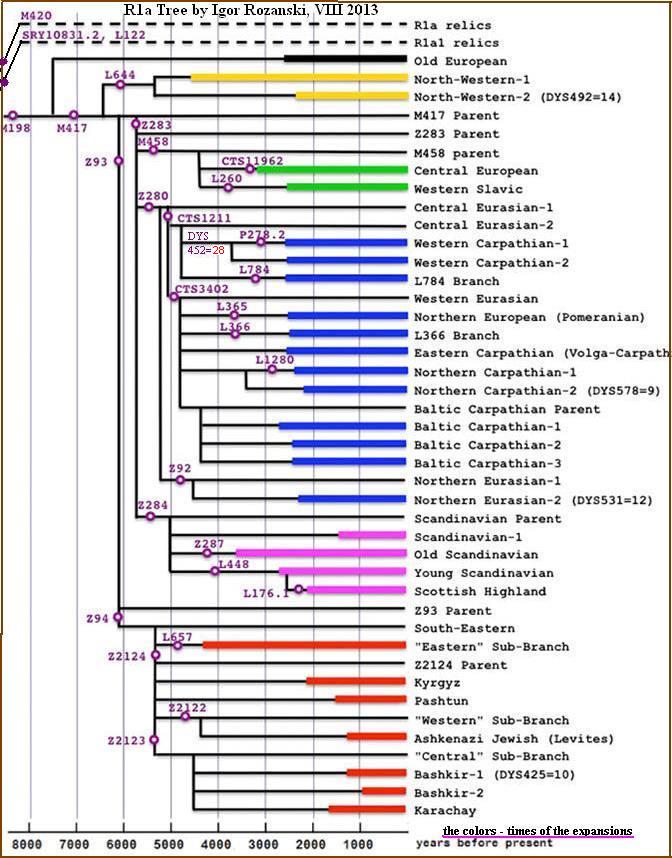
DRZEWO mutacji w rodzie R1a.
Podstawa: ISOGG i FTDNA (Th.Krahn) ora aktualnych danych. Aktualizowano 10.03.2012
DRZEWO mutacji Y-DNA w rodzie R1a. (Aktualizowano 01.08.2014)
Podstawa: YFull experimantal tree, Karafet 2014 oraz aktualnych danych.
Datowwanie YBP oznacza rozwidlenie h.grupy; Lata podano w przybliżeniu
R – 18.400 lat temu M207/UTY2, P224, P227, P229, P232, P280, P285, S4, S8, S9,V45
• R1 – 10.400 lat – M173/P241, M306/S1,P225, P231, P233, P234, P236, P238, P242,P245, P286, P294.
…………………………………………………………………………………..
• • R1a – L62/M513, L63/M511, L145/M449, L146/M420.
• • • R1a1 – L120/M516, L122/M448, M459, Page65/SRY1532.2/SRY10831
• • • • R1a1a – M17, M198
…………………………………………………………………………………….
• • • • • R1a1a1 – M417 – 7.700 lat temu.; mutacja i podział rodu indoeuropejskiego – oddzielenie się CTS4385
• • • • • • R1a1a1a – CTS4385 i L664 – niewielka grupa europejska, północno-zachodnia
• • • • • • R1a1a1b – Z645; kolejny podział rodu indoeuropejskiego na Z283 (Europa) i Z93 (Azja)
……………………………………………………………………………………
• • • • • • • R1a1a1b1 – Z283 – powstanie rodu europejskiego.
• • • • • • • • R1a1a1b1a – Z282 – podział rodu europejskiego na gałęzi M458, Z280 i Y2395.
……………………………………………………………………………………
• • • • • • • • • R1a1a1b1a1 – M458 – Europa Środkowa. (i Zachodnia Ukraina),
• • • • • • • • • • R1a1a1b1a1a – L260 – gałąź zach.-słowiańska: Polska, Czechy, Słowacja.
• • • • • • • • • • R1a1a1b1a1b – CTS11962 – gałąź zachodniosłowiańska.
• • • • • • • • • • • R1a1a1b1a1b1 – L1029
……………………………………………………………………………………
• • • • • • • • • R1a1a1b1a2 – Z280 – duża gałąź bałto-słowiańska;
• • • • • • • • • • R1a- Z92 – gałąź bałtyjska i wschodnia.
• • • • • • • • • • R1a- CTS1211 – gałąź „słowiańska”
• • • • • • • • • • • R1a-YP340 – środkowa Europa
• • • • • • • • • • • • R1a-P278.2 – gałąź „zachodnia karpacka”.
• • • • • • • • • • • • R1a-YP380 – gałąź „stara karpacka”.
• • • • • • • • • • • • • R1a-NN
• • • • • • • • • • • • • R1a-NN
• • • • • • • • • • • R1a-L784
• • • • • • • • • • • R1a-L1280 – północno-wschodnia europejska
• • • • • • • • • • • R1a-CTS3402 – środkowa i wschodnia Europa, gałąź bardzo wsielka.
• • • • • • • • • • • • R1a-Y237 –
• • • • • • • • • • • • • R1a-YP419
• • • • • • • • • • • • • R1a-NN
• • • • • • • • • • • • • R1a-NN
• • • • • • • • • • • • • R1a-YP234
• • • • • • • • • • • • • • R1a-YP295
• • • • • • • • • • • • • • • R1a-YP335
• • • • • • • • • • • • • • • R1a-YPYP346,L366 – Pomorze
• • • • • • • • • • • • • • R1a-L365 – Pomorze, Kaszubia
• • • • • • • • • • • • • • • R1a-NN
• • • • • • • • • • • • • • • R1a-NN
• • • • • • • • • • • • R1a-Y2613
• • • • • • • • • • • • R1a-Y33, CTS8816
…………………………………………………………………………………….
• • • • • • • • • R1a1a1b1a3 – Y2395 i Z284 – gałęzie północno-zachodnioeuropejskie.
• • • • • • • • • • R1a1a1b1a3a – S 5301 i L448 – północna i północno-zachodnia Europa
• • • • • • • • • • • R1a1a1b1a3a1 – CTS4179
• • • • • • • • • • • • R1a1a1b1a3a1a – L176 – Szkocja
• • • • • • • • • • R1a1a1b1a3b – Z287 – starsza gałąź skandynawska
…………………………………………………………………………………………
• • • • • • • R1a1a1b2 – Z93 i Z94 – powstanie gałęzi azjatyckiej i Aszkenazyjczyków
• • • • • • • • • R1a1a1b2a1 – L657 – zwłaszcza indo-irańscy Ariowie
• • • • • • • • • • R1a1a1b2a1a – Y7 – wśród Kazachów i Arabów
• • • • • • • • • R1a1a1b2a1a2 – Z2124 – Eurazja (m.in. wśród Tatarów)
• • • • • • • • • • R1a1a1b2a1a2a – Z2123
• • • • • • • • • • R1a1a1b2a1a2b – Z2122 – głównie żydzi aszkenazyjscy, lewici
• • • • • • • • • • • R1a1a1b2a2b1 – F1345
• • • • • • • • • • • • R1a1a1b2a2b1a – CTS6 – aszkenazyjscy-lewici, z ok. 900 r. po Chr.
• • • • • • • • • • • • R1a1a1b2a2b1b – F2935
• • • • • • • • • • • R1a1a1b2a2b2 – Y57
…………………………………………………. ……………………………………….. ……………………………………….. …………………………………….
• • R1b – M343, itd. – to początek wielkiego drzewa, bratniego dla R1a; zwł. ludność środkowo i zachodnioeuropejska, zwłaszcza italo-celtycka i germańska; w Europie od ok. 2500 lat przed Chr.. Liczna też w Afryce Subsaharyjskiej (np. rejon Czadu i pn. Kamerunu) V88.
…………………………………………………………………………… ……………………………………… …………………………………………………………..
Uwagi
1. Obok symboli wypisano podstawowe mutacje SNP w Y-DNA, określające haplogrupy.
2. Kolorem różowym oznaczyłem własne komentarze. Datowanie zasadniczo w/g http://www.familytreedna.com/public/R1a/default.aspx
3. Nazwy gałęzi zaczerpnięto z PolishProject, R1a1 and Subclades i z drzewa YdnaIP autorstwa I. Rożanskiego, zob. niżej
4. Nie wszystkie firmy, jak np. światowa FTDNA, na bieżąco aktualizują drzewo i wprowadzają zmiany symboli,
stąd różnice i praktyczne nieporozumienia.
Drzewo R1a (z dnia 15.10.2013) – w uproszczeniu
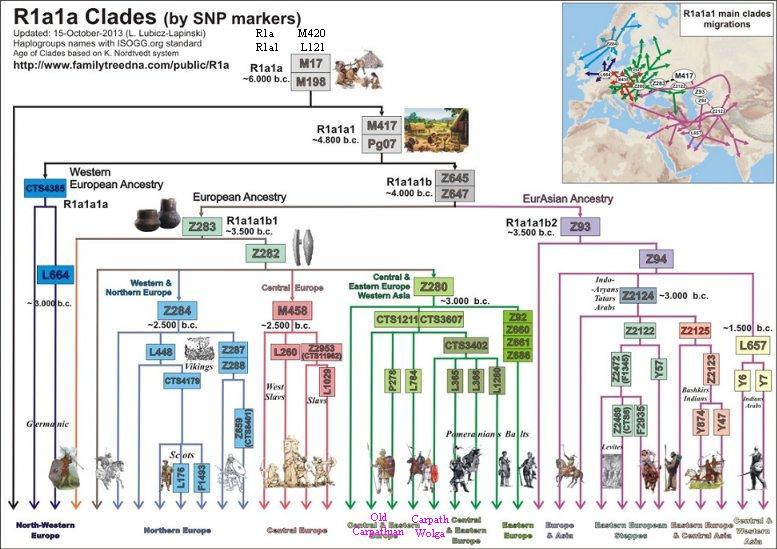
Opis powyższego drzewa
1) Haplogrupy R1a-L62 i R1b-M343 wydzieliły się spod R1 około 15.000 lat przed Chr., według nieostatecznego datowania.
2) Haplogrupa M17 (bez mutacji M417) spotykana jest w ilości śladowej w pasie od Arabii do Wysp Brytyjskich; czas ok.6000 lat przed Chr.
3) Haplogrupa M417 to mutacja ojcowska dla małej grupy północno-zachodniej CTS4385 w Europie i olbrzymiej Z645 euroazjatyckiej. Czas rozwidlenia (split, bifurcation) megahaplogrupy M417 – około 4800 przed Chr., może w Basenie Dunaju.
4) Haplogrupa Z645 około 4000 lat przed Chr. uległa na skutek mutacji Z93 i Z283 oraz migracji ważnemu rozwidleniu na grupę azjatycka i europejską. Rozejście się tych dwóch megagrup Z93 i Z283 z jednej wspólnoty praindoeuropejskiej nastąpiło gdzieś w rejonie Morza Czarnego lub Morza Kaspijskiego.
5) Haplogrupa Z93 dała początek rodu azjatyckiego. To indo-irańscy Ariowie (Indie, Pakistan, Afganistan Kurdystan), Tadżycy, Kirgizi i Żydzi aszkenazyjscy (w Europie).
6) Haplogrupa Z283 to ród europejski. Po kolejnej mutacji, Z282, około 3000 lat przed Chr. uległa rozwidleniu na trzy ważne grupy z mutacjami: Z280, M458 i Z284.
6a) Haplogrupa Z280 jest ogólnie zwana bałto-słowiańską. Jest najliczniejsza z powyższej trójki. Jej obecność rozciąga się zasadniczo od Bałkanów do Bałtyku i od Wołgi do Alp. Czas i przestrzenność haplogrupy Z280 kojarzy sie z rozwojem archeologicznej kultury ceramiki sznurowej (od 1290 lat przed Chr.). W składzie haplogrupy Z280 znajdujemy m.in. grupy karpackie z mutacją DYS452=28, pomorsko-kaszubską L365, wschodniopruską L366, bałtycką i wschodniosłowiańską Z92, północno-wschodnią L1280 itd. .
6b) Haplogrupa M458, zwana środkowoeuropejską; powstała prawdopodobnie na terenie Czech (kultura unietycka?), ale rozciągnęła się przez Polskę do Ukrainy. W tej grupie około 2500 lat przed Chr. powstała podgrupa L260 zwana zachodniosłowiańską, a niekiedy nawet polską.
6c) Haplogrupa Z284, zwana skandynawską, rozprzestrzeniała się głownie na terenie południowej Skandynawii i zachodniej Norwegii (do 30 % miejscowej ludności) oraz w niewielkim procencie na Wysp Brytyjskich i Islandii. Ona zapewne wydajnie uczestniczyła w indoeuropeizacji ludności germańskiej i celtyckiej.

Drzewo rodu R1a od I. Rożanskiego
Sierpień 2013( http://r1a.org/index.htm )
Główne mutacje i czas rozgałęzień i ekspansji. Nazwy przyjęte w środowisku Родство. Ру
Prostopadłe linie wskazują czas rozgałęzienia. Kolory – czas ekspansji (trwałego rozwoju demograficznego).
Ekspansja przestrzenna rodu R1a 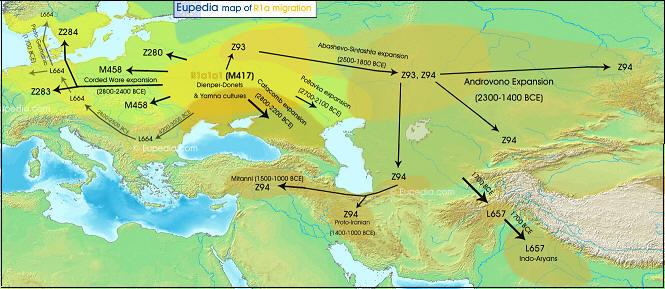
Ekspansja rodu R1a w Eurazji (według portalu Eupedia)
Zadziwiająca jest liczebność, a zarazem rozległość na mapie Eurazji ludności z haplogrupą R1a1a1-M417 z jej podgrupami): od Zachodniej Syberii i Kirgizji na wschodzie, po linię Łaby i wschodnich Alp na zachodzie; od Bałtyku, a nawet wybrzeży Norwegii i zachodniej Szkocji na północy, po Macedonię, Kurdystan, Iran, Afganistan, Pakistan i Indie na południu! Największą częstotliwość tej haplogrupy w Europie, do ponad 60% ludności, stwierdza się w pasie między Bałtykiem a Morzem Czarnym, obejmującym całą Polskę i Zachodnią Ukrainę.
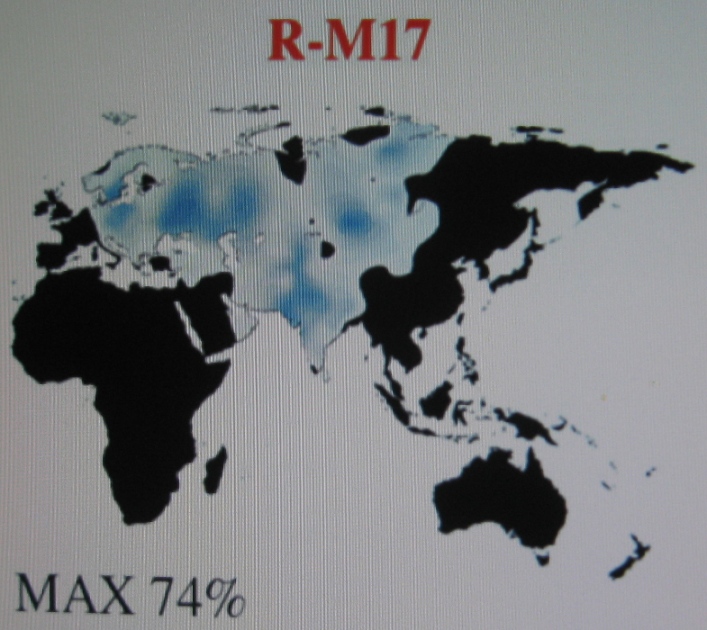 |
Haplogrupa R1a1a1-M417 na mapie Eurazji Widoczne zagęszczenia w rejonie Polski, Ukrainy, Białorusi i Rosji, północnego Pakistanu i Indii oraz Azji Środkowej (Kirgizja). |
Haplogrupą R1a1a1 wyróżniają się, jak widzimy, szeroko rozprzestrzenione na kontynencie europejskim ludy słowiańskie z bałtyjskimi oraz te, które się dowodnie z ich wspólnoty wydzieliły (łącznie z Kirgizami, Tadżykami i indoirańskimi Ariami (o czym niżej). Mając na uwadze to, że spośród kulturowych wyróżników etnosu język jest zasadniczo najbardziej trwałym dobrem, a według niezmiennej zasady, zakłócanej tylko nadzwyczajnymi zdarzeniami,
język otrzymuje się w rodzinie od przodków i praprzodków jako podstawowe dobro tradycji rodowej i plemiennej, więc początek słowiańskiego języka wolno i trzeba łączyć z początkiem nie tylko oddziału R-M458, ale i euroazjatyckiego R-M417, a nadto jego substratu trzeba dopatrywać się już w ojcowskiej haplogrupie R1a (gdzieś w Azji Środkowej), lub nawet jeszcze wcześniej – w haplogrupie R1 (tamże?, Azja Zachodnia?), w której języki satemowe hg R1a łączyły się, jako w jednym źródle, z kentumowymi językami hg R1b i wszystkich jej podgrup (zob. niżej).
Problem pochodzenia Haplogrupy R1a, a zwłaszcza wielkiej podgrupy R1a1a i R1a1a1, jest jednak nadal dyskutowany. Według niedawnej pracy zespołu hinduskich badaczy (ostatnio S.Sharma i zesp., styczeń 2009) uważana do niedawna za ojcowską dla R1a1a haplogrupa R1a1-SRY1532.2 (faktycznie jednak R1a-M420) miałaby się wyłonić w północnych Indiach na terenie Kaszmiru; albo w środkowych Indiach (w plemieniu Saharia) około 18.400 lat temu (licząc 25 lat na pokolenie). Tam wśród plemion Kashmiri Pandits zidentyfikowano 9 osób (4% spośród badanych) z tą haplogrupą, a w plemieniu Madhya Pradesh Saharia 13 osób (23% badanych). Według tych badań, na terenie Kaszmiru lub Indii miałaby powstać także następna mutacja, M17 i M198, dająca początek haplogrupie R1a1a. Dała ona zarazem początek – według tych autorów – hinduskim Indoeuropejczykom, głównej populacji Indii. Obecnie do indoeuropejskiej populacji R1a1a w Indiach należy do 17% hinduskiego społeczeństwa (około 100 milionów ludzi). I co najciekawsze, ta głównie populacji utworzyła najwyższą kastę hinduską, kapłańskich braminów albo przynajmniej nią liczebnie zawładnęła . Przeciętnie blisko połowa, a w głównej ich grupie – nawet 72% kapłanów-braminów wyróżnia się hg R1a1a, podkreślają badacze hinduscy. Daty jakoby sugerują, że z hinduskiej populacji R1a1a miałaby wywodzić się późniejsze populacje Słowian i innych pokrewnych ludów. Autorzy zamieszczają daty tej haplogrupy na Bliskim Wschodzie i w Europie – 11.200 lat temu, a w Azji Centralnej – ok. 8600 lat temu.
Jednak A. KLOSOW, częściowo wspierany badaniami innych genetyków, wskazuje „kolebkę” R1a1a ogólnie w południowej Syberii, w pobliżu Chin. Według Klosowa powstanie mutacji M420, a zatem i haplogrupy R1a, dokonało się gdzieś w okolicach gór Ałtaju, gdzie wśród niektórych plemion, dziś językowo ałtajskich, tureckich i rosyjskich, występuje ojcowska haplogrupa R1-M173 do 20%, czyli w stosunkowo najwyższym zagęszczeniu; choć występuje ona także w Azji Środkowej i na Bliskim Wschodzie – w mniejszym procencie).
Oto przytaczane przez niego fakty:
| Alan Bittles et al. 2007 stwierdził istnienie dość znacznych zagęszczeń R1a1 (M17, M198), czyli obecnie R1a1a, w dwóch północnych prowincjach Chin: Liaoning (północnowschodnie Chiny) i Gansu (północne Chiny), zwłaszcza w plemionach Bonan, Dongxiang, Hui (czyt. Huej) i Salar. W niektórych z nich częstotliwość R1a1a dochodzi do 32 %. Etnolodzy wskazują na istniejące u większości z tych plemion przekonanie o ich obcym pochodzeniu, spoza Chin. Anatole Klyosov2009 w co dopiero opublikowanym Вестник Российской Академии ДНК-генеалогии (Wiadomości Rosyjskiej Akademii genealogii Y-DNA) Том 2, № 5 2009 август, w artykule Древнейшие восточно-азиатские ветви гаплогруппы R1a, (Stare wschodnioazjatyckie gałęzi haplogrupy R1a) dokonał obliczeń i ustalił, że |
R1a1a wywodzi się z południowej Syberii; mutacja M17-M198 powstała tam 20.000 lat temu
bowiem na taki czas wskazują jej haplotypy STR, które Klosov w swojej pracy opublikuje.
Doszedł do wniosku, że jedna część tej populacji z południowej Syberii wyemigrowała przez północnowschodnie i środkowe Chiny do południowych Indii i Pakistanu; w Indiach stanowi starą populację wśród drawidów, ze wspólnymi przodkami z czasu 7000-12000 lat temu.
Druga część, która pozostała „na miejscu” w Syberii, dała później początek być może niektórym starym tamtejszym kulturom archeologicznym, jak np. taszbułatowskiej (8000-7000 lat) i Afanasjewo (rejon Ałtaju 4500-4000 lat temu).
Trzecia zaś część wyemigrowała na zachód, docierając około 12.000 lat temu na Bałkany, by po kilku tysiącach lat około 5000-4000 rozejść się po Europie, w tym także do pontyjsko-kaspijskich stepów, oraz dalej na wschód, za Ural, tworząc kulturę Sintaszta i Andronowo, gdzie uformowała sie populacja Ariów, która w północnych Indiach dała początek młodszej, indoeuropejskiej części R1a1a (o czym niżej).
Datowanie haplogrupy R1a1a na czas około 20.000 lat pociągnie za sobą wcześniejsze datowanie powstania także mutacji M173, czyli haplogrupy R1, datowanej przez T. Karafet et al. 2008 na czas 18.500lat temu.
Klosow jednak dodaje, że konieczne są dalsze ustalenia w temacie datowania i migracji hg. R1 i R1a1a oraz jej rozprzestrzeniania się.
Najnowsza publikacja o starych azjatyckich haplogrupach, w tym R1a1a, zob. Shou et al. 2010 Ocena artykułu w Rodstwo.ru
Klosow, przeprowadzając rewizję dotychczasowego datowania haplogrup europejskich i azjatyckich, w tym także R1a1a, zauważył, że dokonane przez historyków, językoznawców i archeologów datowanie inwazji półwyspu Indyjskiego przez Ariów z północy i zaistnienie tam indoeuropejskiego języka dopiero na połowę drugiego tysiąclecia p.n.e. jest zgodne tylko z poprawnym datowaniem i genealogią hinduskiego haplotypu R1a1a, wyraźnie identycznego z haplotypem wschodniosłowiańskim, lecz nieco młodszego.
Bowiem spośród Prasłowian wschodnich, których wspólny przodek wywędrował około 2750 lat p.n.e na ukraińsko-rosyjskie stepy, część populacji migrowała dalej na wschód, na teren dzisiejszego północnego Kazachstanu i południowego Uralu, tworząc tam rozległą kulturę andronowską (z centrum nad Jeziorem Aralskim). Prawdopodobnie w tej populacji powstała ważna środkowo i południowoazjatycka oraz bliskowschodnia nutacja L432.2, wykryta w lipcu 2011 r.
Niewielka grupa z tej populacji zbudowała sobie około 1800 r. p.n.e m.in. miasto/zamek o nazwie Arkaim na południowym krańcu Uralu. Został on jednak porzucony po około 200 latach. Bowiem po jakiejś katastrofie, którą można kojarzyć z wybuchem (około 1628 lat p.n.e) wulkanu Santorini (zwanym też Thera) na Morzu Egejskim i który pochłonął cywilizacje minojską, a na szerokiej przestrzeni północnej półkuli na długi czas zakłócił pogodę (znaczne ciemności i obniżenie temperatury) ludność Arkaim z R1a1a porzuciła gród i udała sie na południe, zaludniając Kirgizję i Tadżykistan (tam dziś wśród ludności – do kilkadziesiąt procent genetycznych ex-Słowian R1a1a!) oraz północne Indie, a chyba osobno – północny Iran (badania irańskie są dotąd niewystarczające).
Co do hinduskiej populacji R1a1a trzeba wiedzieć i brać pod uwagę, że na podstawie tekstów hinduskiej świętej księgi Rygwedy i innych badań, zwłaszcza archeologii i językoznawstwa, światowa nauka dotąd uważa indoeuropejski lud Ariów, który dał początek kastom braminów hinduskich i hinduskim Indoeuropejczykom, raczej za najeźdźców z północy w połowie II tysiąclecia p.n.e., co potwierdza genealogia genetyczna. Dlatego Genetyka na portalu Eupedia podkreśla: „Kapłańska kasta braminów składa się niemal wyłącznie z haplogrup R1a1, R2 i J2a (chociaż R1a1a stanowi dwie trzecie z linii), przybyłych w ramach indoaryjskiej migracji z zewnątrz w epoce brązu 3500 lat temu”. Genealogia Y-DNA w badaniach A.K. zdaje się potwierdzać pochodzenie językowo indoeuropejskich Ariów od Słowian wschodnich, o czym niżej, w rozdziale o ekspansji Słowian.
|
Uwaga. Obecnie już wiadomo, że w trakcie migracji ludności z haplogrupą R-M417 i jej synowskiej R-Z645 z Europy (Środkowej?) ku Azji Centralnej i Południowej doszło do powstania synowskiej mutacji Z93, która łatwo odróżnia azjatycką część rodu R1a od Europejskiej i wyraźnie wskazuje na kierunek migracji – od Europy ku Azji: Kirgizja, Iran, Pakistan i Półwysep Indyjski. |
Kolejne badania A. KLOSOWA
21.11.2009 r. Journal of Gnetic Genealog 5/2 2009. opublikował właśnie ważną dwuczęściową pracę A. Klyosova na temat rozpoznania interesujących nas haplotypów w haplogrupie R1a1a (teraz R1a1a1), datowania pierwszych wspólnych przodków i migracji. Odnotowuje też i krótko omawia rezultaty badań Underhilla et al. 2009.
DNA Genealogy, Mutation Rates…, cz. I
DNA Genealogy, Mutation Rates…, cz. II
Badania te należą niejako już do historii, gdyż odkrycia nowych mutacji w 2011 r. wprowadziły daleko idące poprawki. Uczciwość naukowa każe jednak je tu zaprezentować.
W części 1. autor omawia swoje sposoby rozróżniania haplotypów i trzy metody obliczania czasu poszczególnych haplotypów od wspólnego przodka (TMRCA); stosowane razem, sprawdzają się wzajemnie i dają znaczną dokładność i pewność.
W części 2. znajdują się praktyczne zastosowania tych metod do obliczania czasu głównych populacji haplogrupy R-M17 (teraz R-M417), bez szczegółowego zajmowania się dopiero co wykrytą podgrupą R1a1a7 (teraz R1a1a1g) oraz innymi wybranymi haplogrupami i populacjami
|
Oto haplogrupa R1a1a – wg. Anatole Klyosov 2009 AZJA EUROPA R1a1a7 (M458) w Polsce może być, wg Klosowa, owocem ponownego zaludniania tego regionu ze wschodu po znacznym wyludnieniu związanym z inwazją populacji haplogrup R1b1b2 około 3200-2500 |
Inne wybrane haplogrupy: 16.000 lat – Haplogrupa R1b (M343) – geneza tej haplogrupy 2325 lat – C2Maorysi, Polinezja. |
Badania innych autorów
Archeolodzy zaś, np. David Anthony (zob. Dyskusja n.12 i mapy), jako pośrednią praojczyznę Słowian przed ich europejską migracją wskazują teren starej kultury Boh-Dniestr, oraz horyzont Jamna (kurhanowy), a z którymi trzeba jakoś połączyć także kulturę ceramiki sznurowej (Corded Ware). To wszystko na wielkim terenie od Karpat Wschodnich po Ural. Najnowsze badania, ogłoszone przez Underhilla (4.11.2009; zob. niżej, rozdz.6) jednak nie bardzo wskazują na ten wschodni teren, lecz na ziemie polskie. Potwierdza go Klosow i Rożanskij (7.12.2009; zob. niżej, rozdz.6). Jeszcze dokładniej, choć ryzykownie, ten temat rozwiązuje paleokraniometria W. Ryżkowa.
Zadziwiająca jest jednolitość, liczebność, a zarazem rozległość na mapie Eurazji ludności z haplogrupą R1a1a1-M417 (z ich podgrupami): od Uralu na wschodzie, po linię Łaby i wschodnich Alp na zachodzie; od Bałtyku, a nawet wybrzeży Norwegii i zachodniej Szkocji na północy, po Indie, Macedonię i Morze Czarne na południu! Największą częstotliwość tej haplogrupy w Europie, do ponad 60% ludności, stwierdza się w pasie między Bałtykiem a Morzem Czarnym, obejmującym całą Polskę i Ukrainę.

| Haplogrupa R1a1a (M17) na mapie EurazjiWidoczne zagęszczenia w rejonie Polski, Ukrainy i Rosji, północnego Pakistanu i Indii oraz Azji Środkowej (Kirgizja).wg. Chiaroni et al. 2009 |
Haplogrupą R1a1a1 wyróżniają się, jak widzimy, szeroko rozprzestrzenione na kontynencie europejskim ludy słowiańskie z bałtyjskimi oraz te, które się dowodnie z ich wspólnoty wydzieliły (łącznie z Kirgizami, Tadżykami i indoeuropejskimi Ariami/Hindusami, o czym niżej). Mając na uwadze to, że spośród kulturowych wyróżników etnosu język jest najbardziej trwałym dobrem, a według niezmiennej zasady, zakłócanej tylko nadzwyczajnymi zdarzeniami,
język otrzymuje się w rodzinie od przodków i praprzodków jako podstawowe dobro tradycji rodowej i plemiennej,
więc początek słowiańskiego języka wolno i trzeba łączyć z początkiem nie tylko polskiego oddziału R-M458, ale i euroazjatyckiego R-M417, a nadto jego substratu trzeba dopatrywać się w ojcowskiej haplogrupie R1a (gdzieś w Azji Środkowej), a nawet jeszcze wcześniej – w haplogrupie R1 (tamże?, Azja Zachodnia?), w której języki satemowe hg R1a łączyły się, jako w jednym źródle, z kentumowymi językami hg R1b i wszystkich jej podgrup (zob. niżej).
6.
Rozwój europejskiego Rodu R1a1a1
R1a1a1g, R1a1a1g2, R1a1a1i… SŁOWIANIE
Ród R-M417, obecny od dziesiątego tysiąclecia przed Chr. (najpierw w swoich przodkach R-M17) zapewne na Bałkanach, nad Dunajem lub w ogóle w Europie Środkowej, na pewno uczestniczył jakoś w procesach rewolucji neolitycznej od około 5500 lat przed Chrystusem: w powstaniu i upowszechnianiu się produkcji ceramiki, hodowli zwierząt i rolnictwa, którego skutkiem był szybki rozwój demograficzny i migracje ludności. W tym czasie w jedności populacji/rodu R1a1a musiał się dokonać proces kształtowania się języka i etnosu, najpierw o wspólnym charakterze indo-słowiańskim, a po rozdzieleniu się – etnosu prasłowiańskiego.
|
Zapewne w rejonie Europy naddunajskiej około 6000-7000 lat temu w rodzie R-M17 pojawiła się mutacja M417, która dała początek wielkiego rodu euro-azjatyckiego R1a1a1. Ze względu na jej późniejsze, tj. około 1500 roku przed Chr. pojawienie się na terenie Indii i Iranu z językiem pokrewnym prasłowiańskiemu i pod szeroko w starożytnych źródłach znaną nazwą „Ariowie”, grupę R1a-M417 można by nazwać ario-słowiańską lub ogólnie aryjską. |
Zob. umiejscowienie mutacji M417, A+, na nici chromosomu Y: 8573735 kb (z FTDNA)

Z tego też, zapewne naddunajskiego czy karpackioego terenu dokonały się po około 3500 lat przed Chr. migracje w różne rejony Europy, na zachód, północ i wschód. Czas i kierunek wędrówek jest dziś rozpoznawany przez warianty haplotypów w STR w haplogrupie R1a1a1, zakodowane w Y-DNA dzisiejszych mieszkańców Europy lub w znalezionych kopalnych szczątkach praprzodków, choć rzadko (bowiem niektóre nowsze plemiona Prasłowian paliły swoich zmarłych; zresztą nie tylko Prasłowianie). Etapy i kierunki migracji pozwalają też rozpoznać kolejne mutacje SNP, aktualnie wykrywane w Europie i Azji, zwłaszcza przez FTDNA.
Wynikiem tych migracji był udział R1a1a1 w powstaniu rozmaitych kultur archeologicznych nad Dunajem, zapewne np. ceramiki wstęgowej, oraz na północ i wschód od Karpat, np. kręgu ceramiki sznurowej z kulturami grobów jednostkowych i amfor kulistych. http://en.wikipedia.org/wiki/Globular_Amphora_culture
Potem nad Dniestrem, Bohem i Dnieprem dochodzi być może do symbiozy z ostatnią fazą kultury Cucuteni-Trypolie u wschodnich Karpat, od rzek Prut i Seret do Dniepru, a w strefie północnej – z kulturą Bałtów – fatianowską.
Genealogia Y-DNA ustaliła, że przodek dzisiejszych wschodnich Słowian z hg R-M417 żył na ziemiach Ukrainy i Rosji od około 2800-2600 lat przed Chr, Następuje teraz rozwój kultury Prasłowian dalej w kierunku wschodnim, ku Wołdze i Uralowi z jakimś udziałem w rozwoju kurhanowego kręgu kultury jamowej (którą A. Klosov przypisuje populacji R1b1a2). Na stepach pontyjsko-kaspijskich praindoeuropejskim kulturom przypisuje się też udomowienie konia, stosowanie powozu i upowszechnianie hodowli zwierząt domowych (zob. Renfrew, Archeologia i język, PWN 2001, s. 249).
(Uwaga. Nie znaleziono dotąd w rejonie Ukrainy i Rosji oczekiwanych przez naukowców genetycznych śladów starszej ludności R1a1a, zwłaszcza takich, których by można datować na czasy ukraińskiego refugium podczas LGM, którego domyślał się już O.Semino i zesp. 2000, tak jak je znaleziono na Bałkanach. Ukraina nie jest więc pierwotną kolebką wszystkich Słowian. Zwłaszcza nie wolno ich genezy datować tam na czasy niedługo przed V wiekiem po Chr., jak to głoszą niektórzy archeolodzy polscy, tzw. skrajni allochtoniści („dnieprowcy”) np. K. Godłowski, a za nimi także niektórzy genetycy (zob. Dyskusja n.5), chyba wprowadzeni w błąd publikacjami znanego niemieckiego archeologa i rasisty Gustawa Kossiny (+1931). Około VI-VII w. po Chr., a więc już w czasach historycznych, ówcześni Słowianie z Ukrainy swoją reemigracją tylko może nieco zasilili osadnictwo południowych Słowian na Bałkanach, przerzedzone klęskami w wojnach z cesarstwem wschodnim oraz na terenie Polski i krajów sąsiednich).
Dziś na Ukrainie, Białorusi i w Rosji żyje około 50% ludności naszych haplogrup (w niektórych starych miastach do 70%). Genealogia Y-DNA, prowadzona przez A. Klosowa i współpracowników, zidentyfikowała nawet starożytnych założycieli dziewięciu plemion ruskich z czasu około 2000 lat p.n.e i późniejszego, nazwanych imiennie dopiero we wczesnośredniowiecznych zabytkach pisanych, m.in. bliscy nam Wołynianie, Bużanie i Biali Chorwaci (kilka kopalnych szczątków ludzi owego osadnictwa hg R1a1a zidentyfikowano nawet w Krasnojarsku w południowej Syberii).
Nieco starsze niż na Ukrainie i Rosji, bo z około 2800 lat p.n.e., korzenie plemion R1a1a1 znajdują się na terenie Niemiec, a więc ród ów jest tam daleko wcześniejszy niż geneza Germanów, zwłaszcza tam, gdzie dziś nadal żyje skondensowane osadnictwo także słowiańskiej hg R1a1a1g/g2 Serbołużyczan (choć zwykło się je datować dopiero na wiek VII po Chr.!). Potwierdza to np. niedawne, dokonane w 2005 r., słynne odkrycie i genetyczne przebadanie cmentarzyska kilkunastu zmarłych, a wśród nich trzech męskich osób z hg R1a1a w Eulau, 40 km za Lipskiem nad rzeką Soławą (niem. Saale) w dorzeczy Łaby w Saksonii. Owe groby ofiar jakiejś starożytnej przemocy pochodzą z około 2600 lat p.n.e i wyróżniają się świadectwami bogatej kultury życia rodzinnego i małżeństwa egzogamicznego. Plemię to prezentowało kulturowe środowisko tzw. ceramiki sznurowej, kojarzonej powszechnie z Indoeuropejczykami. Archeologiczny haplotyp mężczyzny z Eulau genetycy kojarzą obecnie ze staroeuropejską lub skandynawską gałęzią, wydzieloną wcześniej z ogólnego pnia.
Godni uwagi są Nadłabscy Serbowie. jest możliwe, że w jakimś sensie oni, zanotowani przez tzw. geografa bawarskiego jako Zeriuani (Serbianie?), byli tymi, „z których wszystkie plemiona słowiańskie powstały i ród swój – jak zapewniają – wywodzą”. Wśród ludności niemieckiej do dziś około 10% wykazuje haplogrupę R1a1a1, których wspólny przodek żył tam już może około 2700 lat p.n.e. Wyizolowani w niemieckiej społeczności Serbołużyczanie słyną obecnie z tego, ze jest u nich największe w Europie zagęszczenie hg. R1a1a1 (63%). Ale chyba niesłusznie, gdyż jeszcze większe zgęszczenie zostało zauważone na Polskim Spiszu (73%, zob.).
Z rodem R1a1a1 można ośmielić się kojarzyć powstanie i rozwój środkowoeuropejskiej kultury amfor kulistych (http://en.wikipedia.org/wiki/Globular_Amphora_culture), istniejącej w czasie 3400-2800 przed Chr. z centrum na terenie Polski, która około 2900 lat przed Chr. Przeobraziła się ona w rozległą kulturę ceramiki sznurowej, w tym kulturę grobów jednostkowych i toporów gładzonych (http://en.wikipedia.org/wiki/Corded_Ware_culture). Jej południowy region rozciągał się od Małopolski przez Śląsk i Czechy po Solawę (dopływ Łaby), z Serbołużyckim Eulau. Północny region tej kultury przylegał do Bałtyku i rozciągał się od Kujaw i dolnej Wisły, przez Wielkopolskę, rejon dolnej Odry i Meklemburgię po Dolną Łabę (z tym horyzontem można kojarzyć późniejsze R1a1a w jaskini Lichtenesteinhohle z ok. 1000 przed Chr.), zob. Dyskusja, n.16. Kultury amfor kulistych i ceramiki sznurowej powszechnie uważane są za pierwsze indoeuropejskie kultury w Europie. Także słynna kultura łużycka może być w znacznym stopniu przypisana prasłowiańskiemu rodowi R1a1a1.
Populacje haplogrupy R1a1a1 i jej podgrupy żyją dziś w Polsce w największym w całej Europie jednorodnym zagęszczeniu, bo około 57% ludności Polski.
|
Na tablicy rozmieszczenie haplogrup w dzisiejszych krajach europejskich; zob. R1a1a1 w czwartej kolumnie. |
Podobnie jak w Polsce, R1a1a1 (bez uwzględnienia podziału na jej podgrupy) pojawia się w Czechach (dziś 30%), w Słowacji (40%), na Węgrzech (32%; poprzednia błędna liczba: 50%), w Austrii – dziś ok. 25% i w Anglii (5% mieszkańców), Irlandii (dziś m.in. klany Donaldów i Douglasów), Szkocji; Norwegii (do 30% mieszkańców części zachodniej). Stąd właśnie osadnictwo R1a1a1 emigrowało na Wyspy Brytyjskie i do Islandii (dziś tam 23% ludności). Nadto na Litwie, Łotwie i Estonii dziś odpowiednio 38, 40 i 32%.
W sumie, na mapie geno-geografii Europy największe zagęszczenie słowiańskiej (czy byłej słowiańskiej) hg R1a1a1 ciągnie się szerokim pasem od Bałtyku po Morze Czarne (co jest wodą na młyn tym, którym marzy się Polska „od morza do morza”!).
|
W roku 2011, dzięki badaniom Projektu 1000-Genomów, zidentyfikowano wiele nowych mutacji SNP, określanych jako grupa Z albo „zetki”. Jeszcze nie wszystkie z nich dostępne są w powszechnym testowaniu. |
6a. Drzewo genealogiczne
europejskiego oddziału R1a1a1.
Wszystkie mutacje SNP lub STR rozgałęziają drzewo genealogiczne Y-DNA. Mutacje SNP, jako zasadniczo jednorazowe w dziejach człowieka współczesnego, dają początek podstawowym, większym gałęziom drzewa genealogicznego. Mutacje STR, jako szybkozmienne i równocześnie dwukierunkowe („do przodu” i „do tyłu” – powrotne), wytwarzają gałęzie drobne i krótkie. Ale w niektórych markerach STR dotąd doszło tylko do jednej, dwóch lub trzech mutacji – i wtedy są równorzędne mutacjom SNP (niektóre z nich również pojawiają się też dwu lub trzykrotnie). Jedne więc i drugie, SNP i STR, mogą być użyte do budowy drzewa genealogicznego Y-DNA.
Podstawa naukowa i źródła
1. I. Rożanskij i A. Klosow, listopad 2009
– kompendium aktualnej wiedzy o europejskiej części populacji R1a,
(gałęzie przed rozpoznaniem mutacji M458 i haplogrupy R1a1a7)
„Haplogrupa R1a: haplotypy, linie genealogiczne, historia, geografia”
И. Рожанский и А. Клёсов, Гаплогруппа R1a: гаплотипы, генеалогические линии, история, география,
w: Вестник Российской Академии ДНК-генеалогии t.2, nr.6 (listopad), 2009 r., s. 974-1099.
2. A. Klosow i I. Rożanskij, grudzień 2009
o haplogrupie R1a1a1g, czyli gałęzi środkowoeuropejskiej i zachodniosłowiańskiej
„Podgrupa R1a1a1g – M458 – populacje, geografia, historia”
w: Вестник Российской Академии ДНК-генеалогии t.2, nr 7 (grudzień), 2009 r., s.1200-1216.
w języku rosyjskim, gruntowny komentarz do pracy Underhilla et al. 2009 (zob. niżej) w temacie haplogrupy R1a1a1g-M458, uzupełnienie pracy Rożanskiego-Klosowa
„Haplogrupa R1a: haplotypy, linie genealogiczne, historia, geografia” Вестник t.2, nr 6, 2009 r. (zob. wyżej).
3. I. Rożanskij i A. Klosow, styczeń 2010
o bliższym genealogicznym związku gałęzi skandynawskich z kirgiską.
„Migracje z południowej Syberii i Azji Środkowej z punktu widzenia genealogii Y-DNA”
Вестник Российской Академии ДНК-генеалогии, tom 3, nr 1, 2010 r.
4. I. Rożanskij – 2009-2010
kilkanaście map rozmieszczenia gałęzi haplotypów R1a1a1
Google-mapy;
5. P. Szwarew, Forum Rodstvo.ru, omówienie map i drzew genealogicznych
Гаплогруппа R1a: гаплотипы, генеалогические линии, история, география, И. Рожанский и А. Клёсов
6. I Rożanskij, A. Klosow, styczeń 2010
„Migracje z południowej Syberii i Azji Środkowej do północnej Europy z punktu widzenia genealogii Y-DNA”
Миграции из южной Сибири и Средней Азии в северную Европу с точки зрения ДНК-генеалогии
w: Вестник Российской Академии ДНК-генеалогии Том 3, № 1 2010 январь
http://www.lulu.com/items/volume_67/8049000/8049755/2/print/8049755.pdf
Prace rosyjskich genealogów Y-DNA
Analizą mutacji SNP i STR, czyli haplogrup i haplotypów, oraz budową drzewa filogenetycznego i opisywaniem jego gałęzi zajmują się głównie rosyjsko-amerykańscy naukowcy. I. ROŻANSKI i A. KLOSOW w swoich obszernych publikacjach dokonują uporządkowania i genealogicznego opracowania obecnej wiedzy o populacjach haplogrupy R1a i jej podgrup oraz ich historii w Europie i Eurazji. Autorzy mają aktualnie do dyspozycji ponad 2000 haplotypów z bazy Ysearch i geograficznych projektów FTDNA oraz z laboratoriów rosyjskich i prywatnych; najwięcej 67-markerowych.
Praca P. Underhilla (Uniwesytet Stanford)
Publikacja Underhill et al. (publikacja 4.11.2009), znacznie zmieniła dane wcześniejszych autorów, wskazały na okolice Polski jako na praojczyznę Słowian i miejsce powstanie polskiej mutacji R-M458 dla słowiańskiej populacji Europy. Oto niektóre dane polskie i wybranych krajów.
Czas wspólnego przodka podano w tysiącach lat temu, obliczony jest według stawek Zhivotovsky’ego, a więc chyba niewłaściwie obliczonych i dwu-trzykrotnie zawyżonych.
 |
Na lewo: mapa zagęszczenia, czasu haplogrupy R1a1a (M17) i drzewo genealogiczne grupy i podgrup. Na prawo: mapa zagęszczenia, datowania i związku z kulturami archeologicznymi populacji R1a1a1g (M458) – według: Underhill et al. 2009. |
 |
http://www.scribd.com/doc/23322591/Underhill-Et-Al-2009-
Separating-the-Post-Glacial-Coancestry-of-European-and-Asian-Y-Chromosomes-Within-Hap-Lo-Group-R1a
Oto kilka danych Underhilla odnośnie (niestety zawyżonego) datowania populacji R1a1a-M17 i R1a1a1g -M458 w kilku krajach: Polska – lat 11300 i 10700; Słowacja 11200 i 8300; Czechy – 5700; Niemcy 9900 i 7500.
Rosja europejska 8700 i 8000; Ukraina 7400 i 4700; Indie 14000; Pakistan 15000.
Późniejsze wykrycie w tej słowiańskiej podgrupie kolejnej mutacji, oznaczonej jako L260 (R1a1a1g2) i nazwanej P („polska”) jeszcze bardziej związało początki Słowian z tym rejonem Europy.
Prace polsko-amerykańskich genealogów P. Gwozdzia i L. Mayki
W tych pracach Rosjanom dotrzymują kroku, a niekiedy przewodzą polsko-amerykańscy genealodzy – współpracownicy FTDNA: Peter GWOZDZ (USA) i Lawrence MAYKA (USA), którzy zajmują się genealogią wszystkich haplogrup (a nie tylko R1a), ale jakoś związanych z Polską.
L. Mayka prowadzi program FTDNA Polish Project . Rejestruje tam się wyniki analiz przeprowadzonych w pracowniach FamilyTreeDNA dotyczących materiałów z DNA, dostarczonych przez Polaków; internetowa strona przedstawia rezultaty badań i identyfikacje haplogrup Polaków, a w haplogrupie R1a1a1g – podział na klastry, w zależności od haplotypów STR.
Dokładnym rozpracowaniem polskich haplotypów z haplogrup R1a1a i R1a1a1g zajmuje się P. Gwozdz. Na swojej internetowej stronie Polish clades prowadzi analizę haplotypów, grupuje, oblicza czas populacji i ich migracji. W opublikowanej w JOGG Journal of Gnetic Genealog 5/2 2009 jego pracy i pojawia się więcej informacji na temat metody pracy i jej rezultatów (część II). Czas wspólnego przodka haplotypu P .
Prace Łukasza Łapińskiego.
Całością haplogrupy R1a jej mutacji i podgrup w świecie oraz podziałem na genetyczne gałęzie zajmuje się od czerwca 2011 r. genealog Łukasz Łapiński na stronie R1a and Subclades Y-DNA Project.
Natomiast podziałem tego rodu na poszczególne kraje lub większe grupy narodowościowe i regionalne zajmuje się strona Cyndi(Kanada): R1a Y-chromosome Haplogroup Project.
Filogenetyczne drzewa haplogrupy R1a
Autorzy: Anatole Klyosov, Igor Rozhansky, Pavel Shvarev, 2010-2011;
źródło: International Y-DNA Project (w skrócie: YdnaIP) z uwzględnieniem świeżo odkrytych mutacji „grupy” Z.
|
(kliknij) Dostęp do wszystkich map: Gałęzie rodu R1a1a Uwaga. Autorzy drzewa i map aktualizują je często. |
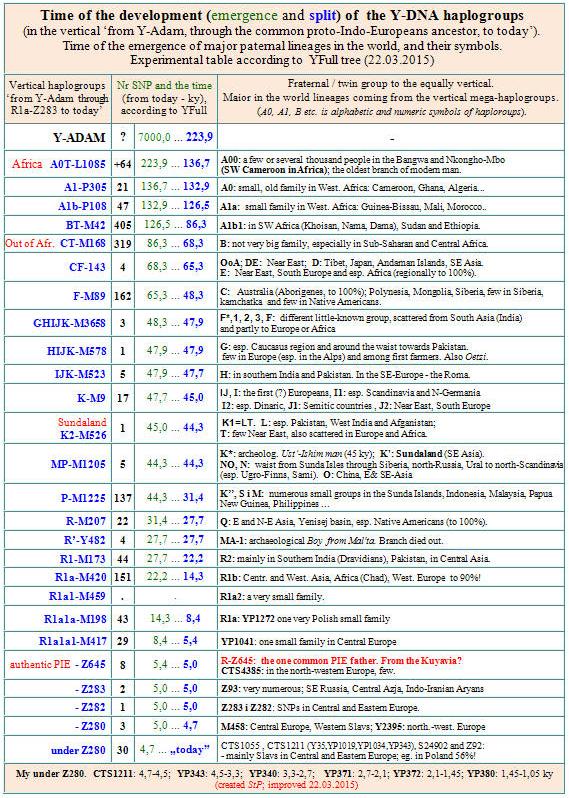
NAZWY GAŁĘZI (polskie i angielskie)
1. Północnoeuropejska – L365, pomorska, z mutacją Z280 2. Środkowoeuropejska – R1a1a1g-M458 3. Zachodniosłowiańska – R1a1a1g2-L260 4. Południowo-wschodnia eurazyjska 4a. Aszkenazyjscy żydzi – z mutacjami Z93 i L342.2; 5. Centralna eurazyjska-3 – zielone L366 i niebieskie (resztkowe) 6. Pólnocna eurazyjska – z mutacją Z280 7. Zachodniokarpacka – P278.2; z mutacją Z280 8. Мłodsza skandynawska – L448, wszystkie, z mutacją Z284 9. Centralna eurazyjska-1 – żółta,. pierwotna indoeuropejska, 10. Północnokarpacka – z mutacją Z280 11. Bałtycko-karpacka – z mutacją Z280 12. Centralna eurazyjska-2, podgałąź czerwona, z mutacją Z280 14. Zachodnia eurazyjska – z mutacją Z280 14a. Zachodnia eurazyjska-2 („wschodniokarpacka”) z mut. Z280 15. Starsza skandynawska – z mutacją Z284 16. Północnozachodnia – DYS388=10, „dziesiętniki” 17. Stare europejskie: R1a, R1a1 i R1a1a (DYS392=13) |
|
Największa rodzina, oznaczona szarą linią ciągłą – to gałąź centralna eurazyjska. |
|


{kind=link}
{kind=link}
{kind=link}
{kind=link}
 |
 |
|
Przykład drzewa genealogicznego |
Kolejne wyobrażenie drzewa genealogicznego i połączeń rodowych R1a1a1. Na końcówkach promieni kolejne liczby tych, którzy zostali przetestowani. Odległość od centrum oznacza liczbę mutacji. Połączenia miedzy gałęziami ujawnia tylko specjalny program. http://s52.radikal.ru/i137/1009/da/3d9d8ca19f49.jpg |
{kind=link}
Podział, datowanie i mapy gałęzi
europejskich populacji haplogrupy R1a i jej podgrup
(wszystkie mapy zob.: Gałęzie rodu R1a1a – International Y-DNA Project; zob. też w forum Rodstwo.ru).
Nowe drzewo R1a – według Igora Rożanskiego.
Stan wiedzy na dzień 15.03.2012 r., http://r1a.org/index.htm
Mutacje, ich powiązanie oraz nowe datowanie;
nazewnictwo wg. ośrodka Russian Academy of Y-DNA Genealogy.

Jest to drzewo, opracowane po rozpoznaniu kolejnych węzłowych mutacji, jak Z283, Z280, Z284, a zwłaszcza „azjatyckiej” Z93 i jej podgrupy L342.2, która wyraźnie oddziela od europejskich R1a1a1 wielkie populacje R1a1a1 na Bliskim Wschodzie, Azji Centralnej, Półwyspie Arabskim i Półwyspie Indyjskim.
Zwróćmy uwagę na dystanse czasowe między oddzieleniem się gałęzi od wspólnego przodka („pnia” drzewa) i powstaniem nowych mutacji, a czasem wspólnego przodka (MRCA) dzisiejszej populacji tej gałęzi i jej ekspansji terytorialnej. Tę odległość między oddzieleniem się gałęzi a MRCA nazywamy demograficzną szyjką butelki.
Szara elipsa wskazuje na połączenie czasu ekspansji rodów R1a1a1 i czasu ekspansji języków indoeuropejskich.
………………….
Aktualne procentowe rozmieszczenie czterech podstawowych mutacji (rodów) haplogrupy R1a w Europie – według diagramu Igora Rożanskiego

(źródło: http://www.rodstvo.ru/forum/index.php?showtopic=1312&st=740)
Drzewo filogenetyczne haplogrupy R1a, aktualny stan wiedzy
według Łukasza Łapińskiego, w projekcie „R1a1a and Subclades”

Zob. w projekcie „R1a1a and Subclades” (Łukasz Łapiński):
http://www.familytreedna.com/public/R1a/default.aspx?section=results
oraz http://img24.imageshack.us/img24/9622/r1acladessnp.jpg
{kind=link}
* * * *
Najstarsze haplotypy z haplogrupy R1a w Europie zostały w 2008 r. rozpoznane przez A. Klosowa wśród ludności niektórych krajów bałkańskich: Serbia, Bośnia, Kosowo i Macedonia, w materiale, który zebrali i opublikowali M. Perićić i L.Barać. A. Klosow datuje je na około dziesiąte tysiąclecie przed Chr. Niesprzyjające warunki bytowe (tzw. szyjka butelki) lub dryf populacji zdziesiątkowały ich liczebność i nie pozwoliły im na pełny rozwój demograficzny. Zanim jednak do tego kryzysu doszło, około polowy trzeciego tysiąclecia przed Chr., owe populacje haplogrup R1a1a miały zapewne wydatny wpływ na rozwój archeologicznych kultur naddunajskich, środkowoeuropejskich i dalszych. Archeogenetyka zapewne będzie miała szansę wypowiedzieć się na ten temat, jak to się stało już odnośnie szkieletów z Eulau, Lichtenstein-Hohle, Krasnojarsk itd.
Zdaniem jednak wielu genealogów genetycznych ów materiał „bałkański”, choć zebrany przez solidne ośrodki naukowe, nie jest w pełni wiarygodny.
Zespół genetyków P. Underhill et al. (listopad 2009, zob. niżej) wykrył zaś ważną europejską mutację R-M458, tworzącą nową haplogrupę R1a1a1g, datując ją, z grubą przesadą, na dziewiąte tysiąclecie przed Chr. i (dość trafnie) wskazując na rejon Polski jakby na teren jej powstania i jakby na praojczyznę znacznej populacji Słowian środkowoeuropejskich, zwłaszcza polskich. Kolejno, po wykryciu w haplogrupie R1a1a1g mutacji L260 ustalono haplogrupę R1a1a1g2, która definiuje wśród Słowian gałąź „polską”, a raczej zachodniosłowiańską.
Zaś szczególną zasługą genealoga genetycznego L. Mayki (USA) jest zidentyfikowanie i zorganizowanie testowania ważnej mutacji „azjatyckiej”, czyli L342.2, która w olbrzymim rodzie R1a1a1(M417) wydzieliła populacje środkowo i południowoazjatyckie oraz żydów-lewitów aszkenazyjskich.
Omówienie głównych gałęzi drzewa haplogrupy R1a
A. Gałęzie rodu R1a przed powstaniem mutacji M417.
Trzy gałęzie stare europejskie (zob. mapa Stara europejska gałąź R1a1a)
To bardzo nieliczni i rozproszeni przedstawiciele trzech najwcześniejszych mutacji w rodzie R1a.
Pierwsza – z mutacją M420, czyli haplogrupa R1a, datowana w Europie na około 9000 lat, czyli 7000 lat przed Chr.
Druga – z mutacją SRY10831.2, czyli haplogrupa R1a1, datowana na około 7000-6000 lat przed Chr.
Trzecia – z mutacją M198 (i M17), ale jeszcze bez mutacji M417, wyróżniająca się markerem DYS392=13 (tu inne gałęzie mają przeważnie 11), czyli haplogrupa R1a1a, datowana na około 4800 lat (jednak najbliższy wspólny przodek, MRCA, żyjącej dziś najstarszej populacji spośród tych trzech haplogrup datowany jest zaledwie na 600 lat przed Chr.).
Niesprzyjające warunki bytowe (tzw. szyjka butelki) lub dryf populacji zdziesiątkowały ich liczebność i nie pozwoliły im na pełny rozwój demograficzny. Zanim jednak do tego kryzysu doszło, około polowy trzeciego tysiąclecia przed Chr., owe populacje haplogrup R1a1a (w tym L170), R1a1a1b i R1a1a1c miały zapewne wydatny wpływ na rozwój archeologicznych kultur naddunajskich, środkowoeuropejskich i dalszych. Archeogenetyka zapewne będzie miała szansę wypowiedzieć się na ten temat, jak to się stało już odnośnie szkieletów z Eulau, Lichtenstein-Hohle, Krasnojarsk itd.
B. Najwcześniejsze gałęzie rodu R1a z mutacją M417-R1a1a1
1) z mutacją L664, na drzewie filogenetycznym oznaczana jako R1a1a1i.
Gałąź północno-zachodnia, pod ojcowską M417, z wyróżniającym się DYS388=10, czyli tzw. dziesiętniki oraz z młodszą mutacją L664. Wydzieliła się, jak oblicza I. Rożański, około 4200 lat przed Chr. i rozprzestrzeniła się także głównie w północno-zachodniej Europie. Składa się z kilku podgałęzi, mających swoim wspólnych przodków, MRCA: 2200, 800 i 500 lat przed Chr. a ich wspólny przodek żył około 4200 lat przed Chr.
2) z mutacją Z645 (dotąd jeszcze nie testowaną)
W rodzie z mutacją Z645 pojawiły się dwie ważne mutacje: „europejska” Z283 (C.) i „azjatycka” Z93 (D.)
C. Gałęzie „europejskiego” rodu R1a z mutacją Z283-R1a1a1g
1) z mutacją M458 – R1a1a1g1 (od 30.12.2011)
1. Środkowoeuropejska (CE) R1a1a1g1, wyróżnia się własną mutacją –M458, powstałą około 2500 p.n.e. którą odkrył i opisał w roku 2009 P. Underhill (Uniwersytet Stanford). Obecna jest w krajach Europy Środkowej, głównie w trójkącie od Czech do ukraińskiego Zakarpacia i Basenu Dunaju (także Małopolski i Śląska, a nawet Wielkopolski, kraju Polan); jest liczna – ponad 16 milionów mężczyzn w Europie. Wyłoniła się prawdopodobnie na południe od Karpat, zapewne w Basenie Dunaju. Wyróżniają ją rzadkie markery DYS464e-f-g, a zwłaszcza mutacja M-458, wspólna z gałęzią zachodniosłowiańską (zob. niżej). Według mapy I.R. środkowoeuropejska gałąź ma dwie podgałęzie, których wspólny przodek żył około 900 lat p.n.e.
W tej populacji, w dunajsko-karpackiej ojczyźnie tej gałęzi prawdopodobnie zrodził jakiś „kult” cesarza Karola niemieckiego i stąd upowszechnić się ogólnosłowiańskie słowo „król”. Tam też mogła być praojczyzna Polan polskich i naddnieprzańskich (zob. u Nestora). Interesujące jest i to, że ekspansja części tej populacji (zwł. z DYS464e-f-g), zapewne w połowie I tysiąclecia n.e., szła – jak wskazuje drzewo genealogiczne – w kierunku z zachodu na wschód, na Ruską Równinę i ku Morzu Białemu, więc w przeciwnym kierunku niż inne, znane w tradycyjnej literaturze naukowej, migracje w czasie wędrówek ludów – ze wschodu na zachód (Wiestnik, s.1026).
2. Zachodniosłowiańska (ZS) R1a1a1g1b (od 30.12.2011). Wyróżnia się mutacją L260, podporządkowaną mutacji M-458 w gałęzi środkowoeuropejskiej R1a1a1g. Wydzieliła się z niej, według ilustr. I.R. 2400 lat przed Chr. Rozmieszczona jest głównie w Polsce, Czechach, Słowacji i na Rusi Zakarpackiej. Powstała zapewne nieco na północ od Karpat. Stanowią ją populacje głównie w dorzeczach Wisły, Odry i Łaby (Polska, Czechy i Słowacja). Przodek żyjącej dziś populacji gałęzi ZS, według mapy I.R., żył około 700 przed Chr. Obydwie gałęzie dość długo żyły w pewnej od siebie separacji, skoro środkowoeuropejska gałąź w części migrowała na wschód i północ raczej jako sama.
UWAGA. Gałęzie zachodniosłowiańska i środkowoeuropejska, mające wspólną mutację M458 z około 2500 lat przed Chr. (zob. wyżej, gałąź CE), oddzieliły się wcześniej, około 3500 lat przed Chr., od gałęzi centralnej eurazyjskiej (CEA); znalazły się jednak w sytuacji długo trwającej demograficznej „szyjki butelki”, albo znacznie zostały eksterminowane i rozwinęły się ponownie dopiero w pierwszej połowie pierwszego tysiąclecia przed Chr.
(Tak więc ewolucja i datowanie gałęzi zachodniosłowiańkiej (ZS), środkowoeuropejskiej (CE) jak i większości pozostałych z R1a1a1 w tym rejonie Europy oraz kojarzona z nimi paleokraniometria kultur naddunajskich i środkowoeuropejskich (już od kultury ceramiki liniowej rytej do kultury ceramiki sznurowej i późniejszych) nie potwierdzają zapoczątkowanej przez niemieckiego archeologa R. Kossinę tezy skrajnych allochtonistów o nieobecności Słowian na naszych ziemiach przed połową pierwszego tysiąclecia naszej ery i o jedynym tylko kierunku ich ekspansji: znad Dniepru ku Wiśle i Odrze!).
2) z mutacją Z280-R1a1a1g2 (od 30.12.2011).
1. Gałąź zachodniokarpacka. Jej wyróżnikiem jest DYS452=28 oraz mutacja SNP L278.2-R1a1a1g2a. Jest rozprzestrzeniona głównie od Karpat i Sudetów po Meklemburgię w Niemczech. Wydzieliła się od pnia drzewa około 5000 lat temu, a wspólny przodek obecnej populacji, MRCA, żył około 600 lat przed Chr. Można tę gałąź łączyć z archeologiczną populacją „sznurowców” z Eulau z czasu około 2600 lat przed Chr.
2. Pomorska (zwana też północnoeuropejską)
Wyróżnia się własną mutacją L365-R1a1a1g2b. Obecna jest głównie na Pomorzu, zwłaszcza na Kaszubach. Dziejami związana jest prawdopodobnie z gałęzią zachodniokarpacką, sięgającą do Meklemburgii i Dolnego Połabia; wiele śladów słowiańskiego języka tamtejszych Wieletów i Obodrzyców znajdujemy w dialekcie pomorskich Kaszubów. Zapewne świadczą one także o poprzednim rozprzestrzenieniu słowiańskich Pomorzan. Gałąź ta wydzieliła się około 5000 lat temu, choć demograficznie rozwinęła sie dopiero około 900 lat przed Chr. Widocznie jakieś niesprzyjające warunki (demograficzna „szyjka butelki”) nie pozwoliły jej jednak wcześniej rozmnożyć się i rozprzestrzenić.
3. Wschodnioeuropejska, rozpoznawalna po mutacji L366-R1a1a1g2c
4. Północno-wschodnia, rozpoznawalna po mutacji Z92-R1a1a1g2d
Nadto bez wyróżniających się własnych mutacji SNP:
5. Północnokarpacka, rozpoznana głównie na terenie Polski. Jej MRCA datowany jest na 200 lat przed Chr.
6. Bałtycko-karpacka, rozprzestrzeniona jest aż do wybrzeży Adriatyku. Wyróżnia się w niej trzy podgałęzie, ale jeszcze bez rozpoznanych własnych mutacji SNP. Datowane są na około 2100, 700 i 500 lat przed Chr., a ich wspólny przodek żył około 2600 lat przed Chr.
7. Zachodnioeurazyjska, właściwie środkowoeuropejska, to ważna gałąź, zapewne ojcowska dla wschodniokarpackiej. Łącznie ze wschodniokarpacką rozprzestrzeniona jest od zachodniej Rosji i Ukrainy po Brytanię. Przodek starszej jej podgałęzi żył około 2000 lat przed Chr.; młodszej – około czasów Chrystusa. Z niej wydzielono gałąź wschodniokarpacką.
8. Wschodniokarpacka, zwana poprzednio „zachodnia eurazyjska-2”, zagęszczona jest głównie w rejonie Karpat. Jej MRCA można datować na 600 lat przed Chr. W jednej jej podgałęzi około początku naszej ery żył przodek tzw. „klanu karpackiego”, który jak sugeruje I. Rożanski, może być identyfikowany ze znanymi ze źródeł dziewięciowiecznych Wiślanami.
9. Północnoeurazyjska, bez wykrytej dotąd własnej mutacji SNP; młoda, rozpowszechniona zwłaszcza na północny-wschód od Polski: Białoruś, Ukraina i Rosja. Jej MRCA datowany jest na około przełomu ery starej i nowej.
10. Centralnoeurazyjska. To wspólnota trzech podgałęzi, blisko spokrewnionych, powiązanych wspólnym przodkiem z około 2800 lat przed Chr. Rejon powstania – zapewne w Europie Środkowej: Karpaty lub Basen Dunaju. Mimo wyłączeniu z niej całej wielkiej gałęzi azjatyckiej z mutacją Z93, rozciąga się ona nadal od Wysp Brytyjskich, poprzez Europę kontynentalną i rejon tzw. Ruskiej Równiny, do Indii (zob. mapa Rożanskiego, gałęzie rozróżnione kolorami). Związane jest to z genezą językowej wspólnoty indoeuropejskiej. Z jej głównej i najstarszej podgałęzi (oznaczonej na mapie żółtymi pineskami), doszło do mutacji Z93, zapewne w trakcie jej migracji rejonem południowego Uralu (kultura Andronowo ze słynnym „grodem” Arkaim) i dalej na Wschód i Południowy Wschód, zwłaszcza ku Indiom i Iranowi.
Obydwie zjednoczone populacje, jedna z mutacją ojcowską M-417 (gałąź żółta), druga z synowską Z93 (gałąź południowowschodnia – na osobnej mapie) i pod nazwą Ariowie („szlachetni”), może około 1600 roku przed Chr. pojawili się na Subkontynencie Indyjskim i na Wyżynie Irańskiej, dając początek etnosom indoeuropejskim w tamtych rejonach (np. język grupy hindi w Indiach i Pakistanie oraz medyjski i staroperski na terenie dzisiejszego Iranu). Wnieśli tam także aryjski system wierzeń religijnych, których wyrazem są święte księgi Rig-Wedy i ich sanskryt (w Indiach) i księgi Awesty (w Iranie).
W późniejszych wiekach z grupy irańskiej wywodzili się słynni Scytowie i Sarmaci – koczownicze i pasterskie ludy stepów azjatyckich i wschodnioeuropejskich.
Zapewne z rejonu nadwołżańskiej i nadkaspijskiej Rosji dokonała się też ekspansja R-L342.2 ku Kirgizji w Azji Centralnej.
W gałęzi centralnej eurazyjskiej około 1300 lat przed Chr. wyłoniła się także europejska podgałąź czerwona (na mapie I. Rożańskiego), zwana orientacyjnie Jasami/Alanami ze względu na pokrewieństwo i obecność haplotypu węgierskich Jasów, wywodzących się z aryjskich Alanów (Alan=Arian, przy regularnej zmianie r’/l).
Trzecia, niewielka podgałąź (zielona na mapie) centralnej eurazyjskiej wyróżnia sie mutacją L366.
3) z mutacją Z284-R1a1a1g3 (od 30.12.2011)
1. Skandynawska, z mutacją L448-R1a1a1g3a. Jej MRCA datowany jest na około 2100 przed Chr. Wyróżnia ją szczególnie mutacja markera YCAII=19,21, podczas gdy w Europie wynosi ona przeważnie 19-23. Oprócz tego marker DYS565=13. Do tej gałęzi niektórzy dopisują, jako ojcowską, populację z Eulau nad Solawą w Niemczech, datowaną izotopowo na rok około 2600 lat przed Chr., nosicieli przypisywanej Prasłowianom kultury ceramiki sznurowej. Skandynawska gałąź R1a1a zapewne wniosła jakieś elementy języka praindoeuropejskiego (odmiany satemowej) i tworzącego się języka prasłowiańskiego do powstającego etnosu pragermańskiego w Niemczech Północnych i południowej Skandynawii (zob. niżej, w rozdz. 9).
2. Skandynawska – z mutacjami L176.1-R1a1a1g3a1 oraz kolejną L175-R1a1a1g3a1a. Członkowie tej gałęzi migrowali także na Wyspy Brytyjskie i Islandię. MRCA jej dwóch podgałęzi datowane są na lata od 200 przed Chr. do 700 po Chr.
D) Gałęzie „azjatyckiego” rodu R1a z mutacją Z93-R1a1a1h
1. Południowo-wschodnia, azjatycka, odznaczająca się mutacją L342.2-R1a1a1h1. Obecna jest na subkontynencie hinduskim, w Pakistanie, Afganistanie i Iranie, krajach arabskich, na Bliskim Wschodzie i w Azji Centralnej.
2. Kirgiska – licząca do około 70 procent ludności Kirgizji. Gałąź wydzielona około 1200 lat temu
3. Aszkenazyjska, także z mutacją L432.2-R1a1a1ah1, rozpowszechniona zwłaszcza w pasie między wschodnim Bałtykiem a północno-zachodnim brzegiem Morza Czarnego. Ta populacja, ma swojego wspólnego przodka, konwertytę na judaizm z populacji znanych z Biblii Aszkenazów (przyszłych Chazarów?); żył około 900 rok po Chr.
4. Arabsko-hinduska – L657-R1a1a1h1a.
|
GENEZA JĘZYKÓW SATEMOWYCH (hindo-bałto-słowiańskich) Na ojczystym terenie, zapewne gdzieś w rejonie Dunaju lub Karpat, w populacji R1a1a1 dokonała się satemizacja języka (dotąd zapewne kentumowego jak nadal w haplogrupie R1b), równoznaczna z formowaniem się języka praindo-prabałto-prasłowiańskiego. Satemizacja polegała głównie na innowacji, że pierwotnie miękkie k’ u satemowców przeszło w s (u kentumowców tylko utraciło zmiękczenie), stając sie zwykłym k lub c. Wnet potem, od około III tysiąclecia p.n.e. (ale jeszcze przed terytorialną ekspansją Prasłowian), nastąpiło oddzielenie się języka indo-irańskich Ariów (głównie z mutacją Z93), a później oddzielenie się języka bałtyjskiego. |
UWAGA.
W rejonie Europy Środkowej nastąpiło spotkanie ludności hg. R1a1a z ludnością kultury pucharów lejkowatych (zapewne hg. I1 i I2), oraz zainicjowanie i/lub upowszechnianie kilku innych kultur, zwłaszcza kultury amfor kulistych i ceramiki sznurowej (Eulau) z toporami gładzonymi i grobami jednostkowymi. Regres demograficzny, trwający w drugim tysiącleciu przed Chr. w rejonie Europy Środkowej, nie był całkowity i nie oznaczał katastrofy kulturowej, zwłaszcza że na sąsiednich ziemiach dzisiejszej Ukrainy i Rosji Europejskiej tego regresu nie było, co umożliwiło późniejsze wsparcie demograficzne i kulturowe dla ziem rejonu Środkowej Europy.
Postscriptum.
Powyższe badania zdają się nie potwierdzać tezy niektórych archeologów o naddnieprzańskim pochodzeniu Polaków i ich haplogrupy R1a, i to dopiero w V wieku po Chr. Co więcej, wyraźnie zauważa się, że Polska leży jakby w centrum starożytnego osadnictwa słowiańskiego haplogrupy R1a. W Rosji np. o rezultatach pobierania próbek DNA wprost pisze się: Poskrob Ruskiego – znajdziesz Polaka (Поскреби русского – найдешь поляка, Газета.ru14.01.2008) – jak przedtem, podobno niesłusznie, tam się mawiało: Поскребешь русского- найдешь татарина – 'Podrapiesz Ruskiego, znajdziesz Tatara’.
* * * * *
7.
GENEZA JĘZYKÓW INDO-EUROPEJSKICH.
Satem i kentum w świetle genealogii Y-DNA
Dla wstępu. Na początku kwietnia 2011 roku The New Jork Times zrelacjonował ciekawą pracę Q.D. Atkinsona z Nowej Zelandii: O śladach elementów fonetycznych pierwotnego języka człowieka współczesnego w dzisiejszych językach; zakwestionowanie glottochronologii (która ograniczała badanie języków do około 4000-5000 lat wstecz):
Phonemic Diversity Supports a Serial Founder Effect Model of Language Expansion from Africa http://www.nytimes.com/2011/04/15/science/15language.html?_r=2&hp
Zob. relację i dyskusję na blogu Dienekesa:
http://dienekes.blogspot.com/2011/04/origin-of-language-in-southwest-africa.html
Równocześnie publikacja o pochodzeniu języków indoeuropejskich w rejonie Anatolii; ponowne zakwestionowanie dotychczasowej glottochronologii; zob. pracę
Language evolution and human history Gray, Atkinson, Greenhill 2011
oraz relację i dyskusję nad nią na blogu Dienekesa:
http://dienekes.blogspot.com/2011/04/indo-european-origins-neolithic.html.
Ważna praca Ryder-Nicholls z 2010 r. na temat początku j. indoeuropejskich w Anatolii
http://www.ceremade.dauphine.fr/~ryder/DPhil.pdf
oraz o początkach indoeuropejskiej monogamii małżeńskiej u Hetytów w Anatolii, Fortunato 2011:
Reconstructing the history of marriage strategies in indo-European-speaking societies: monogamy and polygyny
zob. relację i dyskusję: http://dienekes.blogspot.com/2011/04/proto-indo-european-monogamy.html
Terytorium Anatolii i południowo-wschodniej Turcji na pograniczach Syrii i Palestyny, ze swymi słynnymi punktami protoneolitycznymi i protomiejskimi, np. Gobekli Tepe i Catal-Hoyuk, przyciągają uwagę archeologów, antropologów, historyków i lingwistów. Najprawdopodobniej była to trasa trwającej wiele tysięcy lat migracji z Azji Środkowej (od Ałtaju?) populacji haplogrup R1a i R1b ku Bałkanom i w ogóle ku Europie. Tak nam od kilku lat podpowiada genealogia Y-DNA.
A oto ilustracja pierwotnego zjawiska, zakonserwowanego w starych i izolowanych społeczeństwach rejonu Kaukazu: równoległa ewolucji genów i języka:

Współzależność Y-DNA z geografią (w 60 %) i z językiem (w 64 %) na terenie Kaukazu
(populacje stare i żyjące w izolacji).
Z pracy: Równoległa ewolucja genów i języków w rejonie Kaukazu, O. Balanowski et al. 2011.
Omówienie na blogach: Dienekes Blog oraz Gene Expression, Razib Khan)
Powyższy schemat ilustruje głoszoną na tych stronach zasadę, że źródeł języka należy szukać przede wszystkim u przodków. Dla indoeuropejskich języków satemowych – u przodków R1a, dla indoeuropejskich kentumowych – u przodkiów R1b.
………………………………………………………………………. ……………………………………………………………………….
Wyraźne podobieństwo języków prawie w całej Europie i głównego języka na Półwyspie Indyjskim skłoniło językoznawców do uznania ich pokrewieństwa i nadania im nazwy „języki indoeuropejskie” (logiczniej, więc bardziej poprawnie, byłoby pisanie rozdzielne: języki indo-europejskie). Te związki językowe są świadectwem istnienia u ich podłoża jednego języka, wytworzonego i używanego przez jedną wspólnotę ludzką, żyjącą w zwartej grupie na stosunkowo niewielkim obszarze. Językowi temu nadano nazwę „pra-indo-europejski”, a twórców i użytkowników tego języka nazwano „Pra-indo-europejczykami” (pisownia w różnych krajach odmienna).
Według dość powszechnie przyjętego w językoznawstwie (choć już zakwestionowanego) pogolądu, prawidło ewolucji języków, czyli glottochronologia, miałoby wskazywać na trzecie tysiąclecie przed Chr. jako na czas istnienia jeszcze jednego i niepodzielonego języka.
Gdyby chodziło o ojczyznę języka i wspólnoty prandoeuropejskiej to analiza najstarszego słownictwa, wspólnego dla wszystkich języków indoeuropejskich, miałaby wskazywać na stepy Europy Środkowej i południowej Ukrainy lub Rosji (choć nie brak uczonych wskazujących na Azję Środkową, Anatolię w Azji Mniejszej lub na Bałkany).
Podział języka praindoeuropejskiego na znane nam dziś języki od krańców Europy po Półwysep Indyjski nastąpił na skutek rozprzestrzeniania się i migracji ludności oraz dokonywanych innowacji językowych w nowym małych grupach, od siebie oddalonych i narażonych na kontakty z zupełnie obcymi językami.
Główny jednak podział na grupę satemową i kentumową miałby być, według wielu językoznawców, rezultatem innowacji w tych językach, które pozostawały w centrum obszaru języków indoeuropejskich (j. satemowe, zwł. słowiańskie i indoirańskie) oraz braku innowacji w tych językach, które z powodu migracji znalazły się na peryferiach tego obszaru (j. kentumowe, zwł. celtyckie, romańskie i germańskie).
1. Cechą pierwotnego języka praindoeuropejskiego była palatalność spółgłosek tylnojęzykowych, czyli zmiękczonych k’, g’, g’h.
2. Po podzieleniu i rozejściu się wspólnoty w jednej jej części doszło do spirantyzacji owych spółgłosek tylnojęzykowych palatalnych, czyli przeszły one w spółgłoski szczelinowe typu ś, sz, s. Na skutek tego np. słowo k’entum przybrało brzmienie satem (j. awestyjski) lub sto (j. słowiańskie) i stąd nazwa tej grupy języków: satemowa.
3. W drugiej zaś części pierwotnej wspólnoty językowej doszło tylko do odmiękczenia (dyspatatalizacji) spółgłosek pierwotnie palatalnych. Na skutek tego np. słowo k’entum 'sto’ przyjęło brzmienie kentum 'sto’ i stąd nazwa tej grupy języków: kentumowa.
…………………………………………………………………………. ………………………………………………………………………..
Do niedawna uważano, że do językowej innowacji w postaci satemizacji doszło w centrum pierwotnego terytorium językowego; a ta innowacja nie dotarła do nadal kentumowej peryferii.
Jednak teraz językoznawstwo ma już doskonalsze narzędzie, umożliwiające dokładniejsze ustalenie warunków i okoliczności powstania wspólnoty języków indoeuropejskich oraz fundamentu podziału. Tym narzędziem jest genealogia Y-DNA. Ona bowiem wskazuje na istnienie konkretnej, biologicznej i pierwotnej wspólnoty praindoeuropejskiej w postaci populacji haplogrupy R1. Obecnie łatwo dostrzec, że językami kentumowymi wyróżniają się przede wszystkim populacje haplogrupy R1b (celtyckie, romańskie i germańskie). Natomiast językami satemowymi wyróżniają się populacje haplogrupy R1a (słowiańskie i indoirańskie). Śladem tego, że odmiana kentumowa była wcześniejszą przed satemową formą języka praindoeuropejskiego jest np. kentumowa postać starożytnego języka tocharskiego w Zachodnich Chinach. Badania genetyczne Li et al. 2010 archeologicznych szczątków siedmiu mężczyzn w Xiaohe (Xinjiang), identyfikowanych, choć niepewnie, z Tocharami, wykazały obecność tylko haplogrupy R1a1a.
Można wnioskować, że do podziału języka praindoeuropejskiego na dwie podstawowe grupy doszło więc dopiero wtedy, gdy synowskie haplogrupy R1a i R1b rozeszły się demograficznie i terytorialnie. Ten podział populacji musiał nastąpić wnet po powstaniu haplogrupy R1b, około 16000 lat temu, skoro nie doszło do znacznego zmieszania się populacji co do Y-DNA.
Określenie więc „Indoeuropejczycy”, a raczej bardziej logicznie: „Indo-Europejczycy”, wskazujące dotąd tylko na geograficzną rozległość pokrewnych języków i ich użytkowników, jest teraz uzasadnione także pierwotną jednością ich języka i wspólnym pochodzeniem biologicznym jego twórców.
Ojczyzny zaistnienia i początkowego rozwoju wspólnoty praindoeuropejskiej trzeba szukać zwłaszcza tam, gdzie pozostaje do dziś największe zagęszczenie ojcowskiej haplogrupy R1-M173. Może to być np. w Azji Środkowej, gdzież w rejonie pasma Ałtaju i pogranicza Południowej Syberii, Mongolii, Chin, Kirgizji i Kazachstanu. Np. obecne zagęszczenie haplogrupy R1*, sięgające do 20% (Derenko et al. 2006) w niektórych starych plemionach, np. Teleuci, Szorowie i Tofalarowie w Południowej Syberii pod Ałtajem (u źródeł rzeki Ob i Jenisej) pod granicą z Chinami i Mongolią, świadczyłoby raczej za tym regionem, choć później językowo dość gruntownie sturkizowanym (zob. też Dyskusja n.10).
Drugim terenem pierwotnej wspólnoty praindoeuropejskiej to rejon największego zagęszczenia R1a-M420. Ta haplogrupa nie zostawiła po sobie liczniejszych populacji i jest bardzo rozproszona. Badania M. Requeiro i zesp. 2006, wskazują na Iran jako miejsce pochodzenia synowskiej hg. R1a1. Tam SRY1532.2 zidentyfikowano ogólnie w 3% badanych, a w północnym Iranie – nawet 15%. Potwierdzałoby to wysunięte przypuszczenie o bliskowschodniej kolebce R1a i R1a1, czy dokładniej – Płaskowyż Irański, jak wskazuje W. Ryżkow na podstawie badań paleokraniometrii.
Trzecim regionem zaistnienia i początkowego rozwoju wspólnoty praindoeuropejskiej może być miejsce powstania i oddzielenia sie haplorupy R1b-M343. Ale gdzie to było? Ostatnio Klosow i ludzie z jego kręgu haplogrupie R1b aż do połowy I tysiąclecia p.n.e. przypisują posługiwanie się językiem aglutacyjnym, prototurkijskim. Dowodem na to mają być języki aglutacyjne w niektórych populacjach z grupy R1b, np. Baskowie, Etruskoqwie (?), plemiona północnokaukaskie – należące do rodziny języków sino-kaukaskich lub dene-kaukaskich (w tym jenisiejskich i dene wśród Indian Północnej Ameryki – według teorii Starostina).
Jednak przeciwko korelacji starożytnych haplogrup chromosomowych Y-DNA R1a i R1b z językami praindoeuropejskimi nie może świadczyć fakt niektórych niekonsekwencji w tej dziedzinie. Np. Baskowie, wyróżniający się dość młoda gałęzią haplogrupą R1b1a2, zatraciły swój pierwotny język indoeuropejski, co mogło się stać przecież w trakcie przebywania w rejonie Kaukazu albo na skutek wyjątkowej inicjatywy strony żeńskiej w pierwotnej małej grupie rodzinnej. Podobnie jak żyjąca w Basenie Dunaju ludność, wyróżniająca się haplogrupami R1a1a, I2a, R1b1b2 i E (V-13), przyjęła język małej grupy wojennej Madziarów (czyli przybyłych ze wschodu Ugrów=Węgrów) z haplogrupą N; a mogło do tego dojść na skutek prężnej polityki kulturowej zdobywców tej ziemi wobec braku jednolitości językowej podbitej ludności wieloplemiennej. Podobnie można wytłumaczyć kształt greckiego języka indoeuropejskiego, użytkowanego głównie przez przybyłe do środowiska europejskiego na Bałkanach populacje haplogrupy E1b1b1 o genezie afrykańskiej; ciekawa jest wersja tego języka, bo bliska indoirańskiemu, ale bez satemizacji. Podobnie język Albańczyków, wyróżniających się głównie haplogrupą R1b1b2, jest satemowy, co jest zapewne spowodowane bardziej pierwotnym osadnictwem słowiańskim w tym rejonie i aktualnym otoczeniem.
|
Rozumna i zgodna z Ockhama zasadą oszczędności jest przestroga, by źródeł dzisiejszych języków indoeuropejskich szukać raczej u ich bliższych i dalszych przodków, którzy tworzą i przekazują potomnym swe języki, a dla pojedynczych i mniejszościowych odchyleń (Baskowie, Etruskowie) szukać innych przyczyn i odrębnych okoliczności. |
Interesujące jest, że język siostrzanej względem R1 haplogrupy R2 w południowych Indiach, drawidyjski, jest uważany za pokrewny językom indoeuropejskim w ramach tzw. grupy języków nostratyckich. Bardziej dogłębne badania mogłyby prajęzyków ludności haplogrupy R2 (w Azji Centralnej i w Indiach) mogłyby okazać sie pożyteczne w tej kwestii.
|
Zasadniczo nie są znane przyczyny rozejścia się dróg migracyjnych krewniaczych haplogrup R1a i R1b, a właściwie ich podgrup, oznaczanych głównie jako R1a1 i R1b1b2, z Azji Centralnej ku Europie. Genetyka, jak już wiemy (zob. wyżej, pkt 1), częściowo zna mechanizmy powstawania mutacji, spontanicznych, a także spowodowanych przyczynami zewnętrznymi, czyli mutagenami, często niebezpiecznymi dla zdrowia, a nawet istnienia całej potomnej populacji. |
W każdym razie wraz z oddaleniem się od siebie tych dwóch populacji, dryfowały także i ich języki, różnicując się wolniej wśród grupy wschodnioeuropejskiej R1a, a bardziej intensywnie wśród zachodnioeuropejskiej grupy R1b, co było uzależnione zwłaszcza od bardzo zróżnicowanych warunków życia i zapewne dłuższego przebywania wśród ludów turkijskich.
Duży wpływ na późniejszy kształt języków południowych, zachodnich i północnych w Europie miały bliskie kontakty i mieszanie się ludności słowiańskiej R1a1a i italoceltyckiej R1b1a2 z wcześniejszą, tzw. staroeuropejską, czyli z populacjami weneckimi haplogrupy I (I1 i I2). W południowej Skandynawii doprowadziło to do kreolizacji języków i powstania języka pragermańskiego. Zapewne też znaczny był wpływ tzw. Staroeuropejczyków na pozostałe języki grupy celtyckiej czy italskiej. Mógł też być jakiś wpływ nosicieli dynarskiej haplogrupy I2a2 na kształt języków słowiańskich. Podobnie można by śledzić wpływ języków nieindoeuropejskich populacji bliskowschodnich J1 i J2, kaukaskiej G2a i północno-wschodnioafrykańskiej E, które do Europy, zwłaszcza południowej, napływały w mezolicie lub na początku neolitu (zob. niżej).
Także w populacji indyjskich Ariów, liczącej dziś ponad 100 milionów ludzi z haplotypem słowiańskim R1a1a (16% społeczeństwa Indii i nieco mniej w Iranie), powstały nowe, daleko idące modyfikacje językowe prasłowiańsko-praindoirańskie. W tym języku, zwanym sanskrytem wedyjskim, ułożono przed trzema tysiącami lat w Pendżabie święte hymny Rygwedy, wzywające bogów do udzielenia pomocy wojowniczym Ariom w walce z lokalnymi wrogami (zapewne plemionami drawidyjskimi). W tych tekstach, wiernie przekazywanych ustnie od tysiącleci do czasu ich spisania około XIV w. po Chr., wyczuwa się reminiscencję dokonanego przez Ariów podboju. Populacja R1a1a w Indiach była najprawdopodobniej twórcą także systemu kastowego; w najwyższej kaście, wśród braminów, występują oni do około 72%!
(Nienaukowa więc, krzywdząca dla innych ludów europejskich, a zarazem bardzo szkodliwa dla szacunku Polaków w Europie, jest teza, szerzona na niektórych portalach www. o tożsamości Indoeuropejczyków tylko ze Słowianami, wywyższanie języka i kultury Słowian, ich siły i rzekomej wojowniczości, z wyraźną nutką szowinizmu i pogardy dla innych narodów, które rzekomo tylko dzięki Słowianom stały się indoeuropejskie. Podobne głosy odzywają sie i w niektórych środowiskach nacjonalistów Rosji czy Ukrainy. Jest to próba zastąpienia głupiego przedwojennego pangermanizmu niemieckiego równie głupim i niebezpiecznym panslawizmem genetycznym).
8.
|
Wypowiedź Pawła Szwarewa, genealoga Y-DNA |
|
|
Павел Шварёв (25.1.2011, 13:07) „… Hе забывайте, что мы, славяне R1a , как выясняется, пришли сюда с запада, то есть со стороны Польши, и добрая половина поляков наши реальные братья. Мы от одних предков, просто одни братья когда-то решили податься на восток, а другие остались дома, через какое-то время дети ушедших и оставшихся начали забывать родство, а потом пришла вражда. Реально эта вражда возникла не из нашей среды, это наши правители делили территории, и воевали с помощью нас за эти территории.” |
Paweł Szwarew (25.1.2011, 13:07). Tłumaczenie. „… Nie zapominajcie, że my, Słowianie R1a, przyszliśmy tu, jak się okazuje, z zachodu, to jest od strony Polski i ponad połowa Polaków to nasi dosłownie bracia. Pochodzimy od tych samych przodków. Po prostu jedni bracia kiedyś postanowili udać się na wschód, a drudzy zostali „w domu”. Po jakimś czasie ich dzieci – i ci emigranci, i ci, co zostali w domu – zaczęli zapominać o pokrewieństwie, a potem przyszła wrogość. Ta wrogość nie powstała wśród nas; to nasze rządy dzieliły terytoria i wojowali z naszą pomocą o te terytoria . |
|
Na forum Rodstvo wypowiedź Igora Rożanskiego, autora drzewa rodu R1a1a |
|
|
Igor1961, (10.02.2011). Igor1961, (22.02.2011). Довольно серьезный аргумент в пользу дунайской прародины славян, на мой взгляд. Припятские болота не проходят, поскольку они изобилуют протобалтской топонимикой, а это не стыкуется с низким процентом носителей субклада R1a1a1g-M458 у балтов. Этот субклад четко коррелирует со славянскими миграциями раннего Средневковья. Если бы миграциe начались из региона, близкого к ареалу балтских языков, это подразумевало бы достаточно длительное соседство протобалтов и протославян с неизбежным размыванием границ ветвей. Это наблюдается для балто-карпатской и западной евразийской-2 ветвей R1a1, а также для южнобалтийской ветви N1c1, но не для R1a1a1g, а также северной европейской ветви R1a1a1i. Похоже, ни те, ни другие не входили в состав протобалтских этносов. Вывод: жили где-то в стороне, без непосредственного контакта.” |
Igor Rożanski (10.02.2011). Tłumaczenie. Igor Rożanski (22.02.2011). Tłumaczenie. To, moim zdaniem, dość poważny argument na rzecz naddunajskiej praojczyzny Słowian. Prypeckie błota odpadają, ponieważ one izolują prabałtycką toponimiką, która nie łączy się nawet z niskim procentem nosicieli subkladu R1a1a1g-M458 u Bałtów. Ten subklad dokładnie koreluje natomiast ze słowiańskimi migracjami wczesnego średniowiecza. Gdyby ta migracja zaczęła sie z regionu bliskiego areału bałtyckich języków, oznaczałoby to wcześniejsze długotrwałe sąsiedztwo Prabałtów i Prasłowian, owocujące rozmywaniem granic tych gałęzi. Można to przypuszczać odnośnie gałęzi bałto-karpackiej i zachodniej eurazyjskiej-2, ale nie odnośnie R1a1a1g, a także północnoeuropejskiej [pomorskiej] R1a1a1i. Z pewnością ani ta, ani tamta nie wchodziły w skład prabałtyckich etnosów. Wniosek: żyły gdzieś dalej, bez bezpośredniego kontaktu. |
|
Igor Rożanski na Rodstvo kolejny raz wyraża swoje stanowisko w sprawie genezy języków indoeuropejskich i słowiańskich. |
|
DYSKUSJA
Pytanie:
Badania takich narodów jak Turcy i Węgrzy właśnie doskonale pokazują oderwanie genów od języka. Weźmy zresztą pod lupę taką Grecję. Ten sam język jest tam od ponad 3 tysięcy lat. A co mamy z Y-DNA? Kompletny misz-masz. (…) Spójrzmy na Basków, też osiadłych. Ogromna dominacja r1b. Zajrzyjmy do Celtów (Irlandia, Walia i Szkocja). Też dominacja r1b. A tymczasem ich języki nie mają nic wspólnego z baskijskim. Ba! Baskijski nie jest nawet językiem indoeuropejskim, a celtyckie są. Gdzie tu jakaś logika?.
Odpowiedź:
Tak, jak najbardziej – jest logika! Już sam fakt, że Archeowieści powołały się tylko na wyjątki, jest potwierdzeniem, że poza nimi zasada jakiegoś związku języka z rodami genetycznymi istnieje.
Jaki to związek? Wprawdzie nie ma bezpośredniego („biologicznego”) związku DNA z językiem, ale Y-DNA tworzy przecież ojcowskie grupy rodowe, a języki tworzyły się kiedyś tylko w pierwotnych ojcowskich wspólnotach rodowych oraz plemiennych. Inaczej mówiąc:
Nie Y-DNA, ale pierwotne rody i plemiona, tworzące się „wokół” Y-DNA, mają związek z językami i na odwrót.
a) Pierwotne wspólnoty rodowe niekiedy uległy rozbiciu przez napływ i wmieszanie się nowych populacji (Grecja, Bałkany – będące na korytarzu migracji Azja-Europa). Język natomiast przez rozmaite czynniki zewnętrzne (także polityczne) w małych grupach podlega procesom ujednolicenia.
b) W społecznościach genowo wymieszanych język może być narzucony przez mniejszościowa elitę (np. Węgrzy), która potem może zostać zdziesiątkowana, zostawiwszy jednak po sobie język na trwałe.
c) O języku małej izolowanej społeczności mogą zadecydować także inne czynniki, np. czynnik żeński. Pierwszy przodek plemienia Basków mógł np. oddać inicjatywę stronie żeńskiej i jej otoczeniu, wziętych z jakiegoś plemienia kaukaskiego. Ale to wszystko tylko – wyjątki!
PIERWSZA I NAJSTARSZA NAZWA SŁOWIAN – SPORZY
Sporzy (= liczni, mnodzy, dzielni)Prokop z Cezarei Palestyńskiej, historyk bizantyjski, napisał około 550 roku o Słowianach: „Dalsze krainy na północ zajmują nieprzeliczone narody Antów [Λνταί] (…). Przedtem Sklawinowie [Σκλαβηνοί] i Antowie [Λνταί] mieli jedno imię; albowiem obydwa narody za dawnych czasów zwano Spori [Σπόρoι]; myślę, że to dlatego, iż w rozproszeniu [σποράδην] po wioskach swoich mieszkali. Z tego powodu zajmują obszerne ziemie; w ich posiadaniu jest bowiem większa część krain z tamtej strony Istru (Dunaju)”.Sam Prokop, jak widzimy, ową starą i już nieużywaną nazwę Spori / Sporzy, nieznając słowiańskiego języka, kojarzy sobie na drodze „etymologii ludowej” z greckim słowem sporaden 'sporadyczny, rzadki’. Nowocześni zaś autorzy, np. J. Dobrowski czy P. Szafarzyk, przeważnie widzą w nim zepsute określenie Serbowie. Ale przecież w językach słowiańskich funkcjonuje słowo sporzy (lb.mn.), oznaczające 'liczni, dzielni, sprawni, szybko rosnący’, od prasłowiańskiego słowa *sporъ (lb. poj) i *spori, (lb.mn) oraz praindo-europejskiego rdzenia *sper; np. łac. -sper, staronord. sparr, staroind. sphira-, o podobnych znaczeniach. Stąd właśnie dzisiejsze sporo, spory, sporzy i przysporzyć oraz staropolskie sporzyć (się) ’powiększyć (się), mnożyć (się)’; np. w XV w.: Bóg sporzy 'przymnaża dobro, błogosławi’. W słowie Sporzy znajdują się podstawowe elementy, odpowiadające dużemu, prężnemu rodowi, plemieniu, ludowi lub narodowi. Dlatego przez tysiąclecia mogło ono funkcjonować jako pierwszy, własnego pochodzenia etnonim Słowian, dla Greków niezrozumiały i dlatego wśród nich już nieużywany – jak zanotował Prokop z Cezarei. |
POSZUKIWANIA KRANIOMETRYCZNE W. Ryżkowa (zob.)
Kraniometria wskazuje na powstanie odrębnego haplotypu jako aryjskiego, identyfikowanego z haplogrupą R1a1a, na południu od M. Kaspijskiego, na terenie Płaskowyżu Irańskiego (np. Hotu-Cave?) datowanego przez K. Coona na około 20000 lat temu. Stamtąd po około 10000 latach nastąpiła migracja przez Anatolię (Catal-Hojuk) i Bałkany (ok. 7000 p.n.e.) ku Środkowej Europie, a około 2350 p.n.e. na Nizinie Rosyjskiej (zwłaszcza Ukraina Zachodnia i basen Wołga-Oka). Kraniotyp aryjski zdecydowanie widoczny jest w kulturach ceramiki wstęgowej rytej, ceramiki wstęgowej kłutej, ceramiki pucharów lejkowatych, kulturze ceramiki sznurowej (stwierdzonej na Podkarpaciu w Szczytnej już sprzed 4000 p.n.e!).
UDZIAŁ SŁOWIAN w TWORZENIU STAROŻYTNYCH
KULTUR Archeologicznych
Populacje haplogrupy R1a z jej podgrupami są wraz z populacjami haplogrupy R1b głównymi twórcami nie tylko języków z rodziny indoeuropejskiej, ale także wielkiego dziedzictwa kultur duchowych i materialnych świata starożytnego i nowożytnego, zwłaszcza Europy.
Ubogi i prymitywny materiał ceramiczny tzw. kultury praskiej, korczakowskiej czy pienkowskiej i podobnych, przez archeologów wzorem Kossiny przypisywany Słowianom jako ich rzekomo pierwszy wytwór, w świetle wiedzy o haplogrupach R1a1a i R1a1a1g staje w zupełnie innym świetle. Ta kulturowa prostota mogła być wytworem tylko niektórych, do V w. zapewne jakoś odseparowanych, niewielkich populacji słowiańskich, poruszonych ówczesnymi migracjami i zmianami demograficznymi; nie może ona więc świadczyć o prymitywizmie całej olbrzymiej populacji R1a1a wcześniej i w połowie pierwszego tysiąclecia po Chr., obecnej przecież w mniejszym lub większym procencie na terytorium od Pirenejów, Islandii, wybrzeży Norwegii (Wikingowie) i Bałkanów – po Ural i dalej na wschód, twórców wielu wielkich, znanych i wyrafinowanych kultur.
Ci właśnie, żyjący na terenach Europy Środkowej, Południowo-Wschodniej i Wschodniej Prasłowianie R1a1a i R1a1a1g (oraz populacje z nich wydzielone) byli na tych ziemiach twórcami lub współtwórcami wielu kultur archeologicznych (jak to ukazują mapy na portalu Eupedii, zob. Dyskusje 18).
|
Główne archeologiczne kultury, Tu trzeba z niewielkimi zmianami powtórzyć to, co wyżej powiedziano o populacji R1b, że zdefiniowaną genetycznie populację R1a trzeba łączyć z satemowym wariantem języka indoeuropejskiego, pewnym zubożeniem form ceramiki na rzecz ceramiki sznurowej, początkiem epoki brązu w Europie, obniżeniem kultury rolnej na rzecz hodowli zwierząt i pasterstwa, przejściem od matriarchatu do patriarchatu, wprowadzeniem grobów jednostkowych i kurhanów nagrobnych, udomowieniem i wykorzystaniem konia oraz rozwojem cech wojennych. Najgłębiej, choć może zbyt ryzykownie, te sprawy traktuje W. Ryżkow, który na podstawie paleokraniometrii przypisuje klanowi R1a1a tworzenie następujących kultur w Europie: k. ceramiki wstęgowej rytej, k. ceramiki wstęgowej kłutej, k. ceramiki pucharów lejkowatych, a zwłaszcza k. ceramiki sznurowej. W kulturze unietyckiej (głównie ziemie Austrii, Czech i Śląska) proporcje udziału klanu aryjskiego R1a1a ważą się względem udziału kultury pucharów dzwonowatych klanu R1b1b2 od 10% do 90% (np. na Śląsku) udziału aryjskiego. Z kultury unietyckiej ewoluowała kultura łużycka. |
Uwzględniwszy dotychczasowe dane antropologii, archeologii i językoznawstwa historycznego, wielu może odczuwać ogromne zaskoczenie dostarczonymi przez genealogię genetyczną, zwłaszcza geno-geografię danymi i powyższymi wnioskami. Ale trzeba też zachować ostrożność, przynajmniej do czasu przebadania ludzkiego materiału archeologicznego oraz udoskonalenia metod datowania, które nadal doprowadzają do wielu rozbieżności pozostawiają wiele niepewności chronologicznych.
W każdym razie teraz od wielu archeologów, zwłaszcza skrajnych słowiańskich allochtonistów, powyższe fakty będą wymagały wiele przysłowiowej wielkoduszności i przyznania się do pomyłek, zrozumiałych przecież z powodu dotychczasowej niewystarczalności narzędzi do ustalania etnicznego i czasowego znakowania kultur.
* * * * *
W nowym i korzystnym dla nas świetle staje także wyjątkowa zwartość i liczebność etniczna oraz tożsamość słowiańska Polaków, obecnych od wielu tysięcy lat na tej ziemi (Underhill ustalił nawet blisko 12 tysięcy lat), choć może z okresami znacznego regresu demograficznego, tzw. pustki osadniczej. Widać to szczególnie na tle sąsiednich krajów, zwłaszcza Niemiec (gdzie Germanów z normańskiej haplogruopy I-1 – nieco ponad 40%, ze zgermanizowanej części celtyckiej hg R1b 40%, a ze słowiańskiej R1a1a i R1a1a1g – 20%) oraz mieszkańców Czech i Słowacji (gdzie oprócz Słowian żyje dość liczna grupa celtycko-germańska z R1b odpowiednio Czechy 28% i Słowacja 17% oraz hg południowoeuropejska I-2, po 18%). I zadziwia nadto, zwłaszcza wobec genetycznej różnorodności w haplogrupie zachodnioeuropejskiej, genetyczna jednolitość całej haplogrupy słowiańskiej R1a1a, w której dotąd nie doszło do żadnej dalszej mutacji i wydzielenia się osobnej podgrupy genowej; na kontynencie europejskim (Polacy, Słowacy, Macedończycy, Ukraińcy, Rosjanie…) różnimy się tylko osobniczymi allelami w STR.
Szkoda tylko, że ci Słowianie, choć tak liczni, nie utworzyli wcześniej, już w starożytności, jednej zwartej grupy politycznej i własnego wielkiego i trwałego państwa. Teraz trzeba ich identyfikować nie tylko z późniejszymi Sklawinami, Antami i Wenedami (odVI w. po Chr.), ale i pod wieloma innymi, wcześniejszymi nazwami ludów starożytnej Europy, może łącznie z niektórymi „Sarmatami” (od III w. przed Chr. po późne średniowiecze), niektórymi „Scytami” (znanymi co najmniej od VII w. przed Chr.), Aorsami, Neurami, Budynami, Melanchlajnami, a zwłaszcza Ariami (ok. 1500 lat przed p.n.e.). Trzeba ich więc widzieć, i to już w starożytności, na Bałkanach i nad Dunajem, na rozległym terenie od Bałtyku po Morze Czarne, od ziem Rusi po Morze Aralskie, a nawet od Uralu po ziemie północnych Indii, Pakistanu i Iranu – wszędzie tworzących niekiedy prymitywne, ale niekiedy i bardzo rozwinięte i wyrafinowane kultury archeologiczne (zob. wyżej i na mapach Eupedii:
http://www.eupedia.com/europe/neolithic_europe_map.shtml) oraz systemy protopaństwowe.
Dzisiejsza przestrzenność i niemal jednolitość języków słowiańskich, nakładająca się na przestrzenność haplogrup R1a, każe nam zakładać podobną przestrzenność wcześniejszego, niezróżnicowanego jeszcze języka prasłowiańskiego. Biorąc jednak pod uwagę fakt, że język jest głównym czynnikiem różnicującym ludzi i wytwarzającym świadomość odrębności etnicznej, ta wielka jednolitość i przestrzenność prajęzyka była zarazem hamulcem procesów politycznych. To tłumaczy trudności w identyfikacji i niewielką obecność Słowian w tekstach starożytnych historyków i geografów. Brak wyraźnego czynnika zagrożeń i między etnicznej rywalizacji nie wpływał na przyspieszenie integracji społecznej i mobilizacji gospodarczej. Dopiero w bliższym zetknięciu się z południowo i zachodnioeuropejskimi, zorganizowanymi społecznościami i państwami Słowianie uświadamiają sobie swoją odrębność, a nawet nadają sobie nazwę etniczną: Słowianie, jako ci mówiący zrozumiałymi słowami, w przeciwieństwie do „niemówiących” (psł. „niem„) lub mówiących niezrozumiałą mową, czyli Niemcami. Nazwą tą określano początkowo zapewne prawie wszystkich Nie-Słowian.
9. Nieindoeuropejskie haplogrupy i poPulacje w Europie,
zwłaszcza przedindo-europejscy Wenetowie / Wenedowie
Kiedy indoeuropejskie populacje R1a i R1b zaczęły w polodowcowym mezolicie, neolicie lub epoce brązu zasiedlać Europę, zastały tu zapewne bardzo rzadkie osadnictwo ludności starszej, przed-indoeuropejskiej, która przeżyła okres wielkiego zlodowacenia. Pod względem Y-DNA to haplogrupa I (I1 i I2).
Tak więc pod względem pochodzenia ojcowskiego ludność Europy nie jest zupełnie jednolita. Populacje pierwotnie indoeuropejskich haplogrup: „słowiańskich” R1a1a1 i „italo-celtyckich” R1b1a2, czyli twórców języka praindoeuropejskiego, stanowią około 70 procent Europejczyków. 20 procent to chyba jedyna przedindoeuropejska haplogrupa I (I1 i I2), czyli prawdopodobnie jacyś „Wenetowie”.
Pozostałe 10 procent to zwłaszcza bliskowschodnie J1 i J2, północnoafrykańsko-egejska E, północnosyberyjska i ugrofińska N, małoazjatycka/kaukaska G i hinduska/cygańska H.

1. HAPLOGRUPA I (I1 i I2) czyli przed-indoeuropejscy Enetowie/Wenetowie/Wenedowie(?).
Ważna publikacja: A.Klosow, Gaplogruppa I, Wiestnik Rossijskoj Akademii DNK-genealogii, 3/1 2010 (s. 96-158)
http–www.lulu.com-items-volume_67-8049000-8049755-2-print-8049755.pdf
Między 48.000-40.000 lat temu pojawiła się w Europie pierwsza populacja z gatunku homo sapiens-sapiens („człowiek współczesny”). Był to tzw. „człowiek kromanioński” (Zachodznia Francja), albo „człowiek bohunicki”, zidentyfikowany w miejscowości Bohunice k.Brna (Czechy) oraz w kilku miejscowościach Europy Południowej (Włochy), Środkowej (Bułgaria, Morawy i Polska-Dzierżysław) i Wschodniej (Ukraina i Rosja), np. „człowiek z Kostienek nad Donem, 30000 lat. Nie znaleziono dotąd ich wyrobów-narzędzi, ale np. przez porównanie z człowiekiem z Ahmarian (Turcja) uznano bohunicjan za ludzi współczesnych, ich kulturę materialną określono jako proto-oryniacką, a jako ojczyznę ich pochodzenia uznano Bliski Wschód. Z powodu wystąpienie około 40.000 lat temu zjawiska „zdarzenie Heinricha4”, spowodowanego chyba wulkanicznym Ignimbrite, którego skutkiem było gwałtowne, trwające kilkaset lat zanieczyszczenie i oziębienie atmosfery, człowiek bohunicki mógł prawie całkowicie wyginąć.
Po tym zdarzeniu, około 38.000 lat temu pojawia się już człowiek „klasycznej” kultury oryniackiej. Kultura ta znana jest w całej Europie 35.000-29.000 lat temu, także w Polsce, np. w jaskini w Obłazowej koło Nowego Targu (zob. Dyskusja n.13). W sumie to około 20% mieszkańców Europy.
Pod względem genetycznym człowiek oryniacki odznaczał się prawdopodobnie haplogrupą IJ, powstałą w Mezopotamii, około 50000 lat temu. Stamtąd, według przypuszczeń A.Klosowa, poprzez Kaukaz dostał się na ziemie nad Wołgą i Donem. Jego śladem może być tzw. człowiek z Kostienek (z około 30.000 lat). Ale nie istnieje ta mutacja do dziś, gdyż jej populacja mogła wyginąć w czasie ostatniego wielkiego zlodowacenia (MGL, od około 20.000 lat temu). Natomiast pozostała po niej synowska populacja z haplogrupą I (M170) oraz jej podgrupy I1 (M253) i I2 (P37.2). Wyłoniły się one już przed okresem wielkiego zlodowacenia, 30.000-20.000 lat temu.
Językowo były to populacje staroeuropejskie, przed-indoeuropejskie. Zapewne one zapoczątkowały i rozwijały od około 28.000 do 22.000 kulturę grawecką (znaną także z jaskini Obłazowej), oraz młodsze jej pochodne, np. polodowcową kulturę magdaleńską, spotykaną także w kilku regionach Polski Południowej. Zapewne ta haplogrupa I, rozproszona po Europie w kilku genetycznych podgrupach, tworzyła populacje, zwane w starożytności i średniowieczu Enetowie/Wenetowie/ Wenedowie (nie mieszać z Wenecjanami!); zob. Dyskusja n.3).
 |
Przedindoeuropejska haplogrupa I i jej podgrupy w Europie, wg. Chiaroni et al. 20091. Skandynawia, Niemcy, Nadrenia: to germańskie hg. I1 (M253) i I2b1 (M223). 2. Sardynia, Płw. Iberyjski, Irlandia: to hg I2a1 (M26). 3. Chorwacja i Ukraina: to dynarska, słowiańska hg I2a2 (M423). |
A) Haplogrupa I1, czyli głównie „Wenetowie/Wenedowie” germańscy.
Z ojcowskiej haplogrupy I gdzieś na południu Europy około 22.000 lat temu, a więc jeszcze przed LGM, wyłoniła się w drodze mutacji M253 haplogrupa I1 (i-jeden); jej populacja przetrwała zlodowacenie zapewne na Rosyjskim Niżu lub w Akwitanii (południowo-zachodnia Francja). Czas jej rozwoju i ekspansji przypada na drugie tysiąclecie pne. Rozprzestrzeniła sie ona później na północ ku Finlandii i na południe – ku Jutlandii i północnym Niemcom. Dziś w Szwecji stanowi 42% ludności, w Norwegii 36%, Islandii 33%, Danii 30%, Finlandii 28%, a w Niemczech ponad 20%.
Udało sie ustalić tzw. pierwszych przodków poszczególnych populacji haplogrupy I1. Wspólny przodek haplogrupy I1 w Anglii, Irlandii i Szkocji żył około 1425 p.n.e; w Skandynawii – 1375; w Europie kontynentalnej od Hiszpanii do Słowacji – 1425; we Europie Wschodniej łącznie z Polską – 1225; tak samo w Niemczech; Bliski Wschód – 1475 p.n.e. Ekspansja tej haplogrupy czasowo kojarzy się więc z ekspansją słowiańskiej haplogrupy R1a1a.
Tu dygresja o powstaniu Germanów. Choć haplogrupa I1 pojawiła się w Skandynawii wcześniej, tworząc tam wielkie starożytne kultury przedneolityczne, to jednak około 1300-1400 lat p.n.e. na Półwyspie Skandynawskim przebywała ich bardzo mała wspólnota. Jeden bowiem mężczyzna tej haplogrupy około 1375 p.n.e. dał tam początek wszystkim późniejszym populacjom nordyckim tej haplogrupy. Nieco później pojawiło się tam także i plemię „słowiańskie” z hg R1a1, oraz „północno-zachodni oddział” I2b1 (M223), a w ramach migracji z celtyckiej kultury Hallstatt pod Alpami ok. 700 p.n.e. dotarły tam plemiona haplogrupy R1b1b2. I właśnie z tych trzech grup: I1+I2b1, R1b1b2 i R1a1a na styku Skandynawii i północnych Niemiec powstali Germanowie. Dziś wśród nich około 40% to haplogrupa I1+I2b, około 40% – R1b1b2 i około 20% – R1a1a. Tak więc rasistowskie myślenie niektórych dawnych Niemców o czystości rasowej Germanów zostało przez dzisiejszą genealogię genetyczną zupełnie skompromitowane.
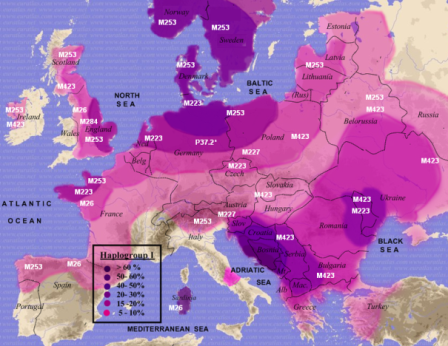 Populacja hg I i I2; główne podgrupy |
Na lewo: |
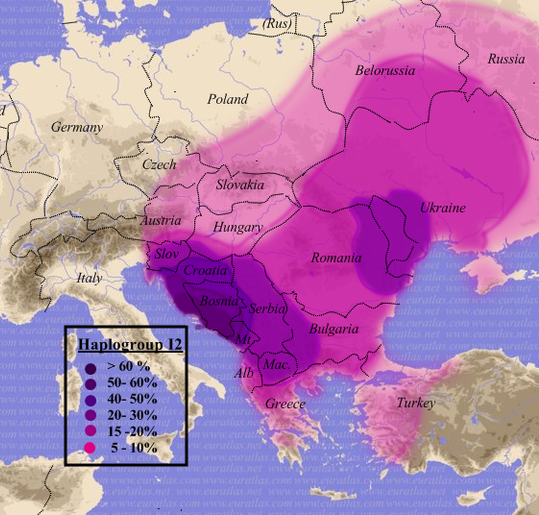 |
B) Haplogrupa I2, I2a1, I2a2 – głównie „Wenetowie/Wenedowie” słowiańscy.
W populacji hg I po mutacji M438, około 21.000 lat temu, a więc także przed ostatnim zlodowaceniem, wyłoniła się haplogrupa I2. a z niej,
Powstała w niej około 20.000 podgrupa I2a1 (M423), która w swoim czasie, wraz ze Słowianami (R1a1a), zdominowała niektóre rejony bałkańskie i środkowo-wschodniej Europy: Chorwacja 42%, Bułgaria 33%, Serbia 27%, Rumunia 17%, Węgry15%, Białoruś 25%, Ukraina 12%. pd.-zach. Rosja 11%, Polska 10%, Słowacja 10%). Nic dziwnego, że w starożytności niektórzy autorzy nie umieli rozróżnić między rdzennymi Słowianami (R1a1) a zeslawizowanymi Wenetami (I1a2); już wtedy obie populacje tworzyły jakby jeden etnos. To zapewne ci Wenetowie tworzyli najstarsze kultury archeologiczne na stepowych terenach Ukrainy i południowej Rosji, np. kurhanową czy ceramiki sznurowej, zanim nie dołączyła do niej młodsza, przybyła z zachodnich Bałkanów czy już znad Dunaju populacja słowiańska R1a1.
Udało się ustalić pierwszych wspólnych przodków dla poszczególnych populacji Haplogrupy I2. Okazała się ona bardziej niż hg. I1 odporna na przeciwności życia w tamtym czasie. I tak: w Anglii wspólny przodek wszystkich populacji I2 żył 13000 p.n.e.; W Irlandii 14200 p.n.e.; w Szkocji 14800 p.n.e.; w Skandynawii 13000 p.n.e.; Europa kontynentalna od Hiszpanii do Słowacji 1425 p.n.e.; Niemcy 9675 p.n.e.; Europa Wschodnia łącznie z Polską 14200 p.n.e..
Tu dygresja o powstaniu Rusi. W ruskich Latopisach napisano: „Przyszli Waregowie. Nazywali się Ruś… Mieli język ruski i słowiański„. W XVIII w. dopisano z fantazji, że Waregowie przyszli zza morza i byli Normanami. Faktycznie to chodziło raczej o plemiona Rugów/Rusów, żyjących wśród Słowian co najmniej od II w. w pasie miedzy Bałtykiem a Morzem Czarnym i zmagających sie z naporem germańskich Gotów. Ich genetyczne haplolgrupy to I1 (populacja bałtycko-skandynawska) oraz I2a2 (populacja południowosłowiańska). Ogólnie wśród dzisiejszych „Ruskich” hg I2a1 to 15%, a I1 – 6,5% ludności, żyjących obok obok R1a1a – 48% i N1c – 14%. Zapewne z hg I1 był Ruryk, Olga, Włodzimierz i inni protoplaści Rusi kijowskiej i nowogrodzkiej.
Zob. o tym: Klosow, Haplotypy południowych i bałtyjskich ruskich Słowian: cztery plemiona? w: Proceedings... t. II/5 (2009), s. 801-815 (por. A. Klosow).
Dygresja o powstaniu Bałtów. Uważa się, że etnos i język Bałtów (czyli zwłaszcza Prusów, Litwinów i Łotyszy) wydzieliły się ze słowiańskich, o czym świadczą niektóre wspólne innowacje językowe bałto-słowiańskie, i to najpóźniej około 1000 lat przed Chr. Dziś w rejonie Prabałtów spotykamy słowiańską haplogrupę R1a1a około 40% i przed-indoeuropejskie: bałtycko-skandynawską I1 i bałkańską I2a2, razem około 6% (Białoruś 25% I2a2). Można więc uważać, że haplogrupy I1 i I2 (Wenetowie bałtyccy) uczestniczyły w tym regionie w ewolucji słowiańskiego języka i wydzieleniu się jego formy bałtyckiej (por. A.Klosov).
Haplogrupie I2a1 (M423), obok hg E1b1b1a2 (V-13), genetycy przypisują upowszechnianie rolnictwa, przynajmniej w południowo-wschodniej Europie, przejętego w okolicach wschodniego wybrzeża Adriatyku od przybyłej z Anatolii haplogrupy J2b2 (M241). Zob. Dyskusja n.11.
Natomiast jej siostrzana podgrupa I2a2 (M26), wyłoniona około 19.000 lat temu, zdominowała Sardynię (39%) oraz nielicznie przetrwała na północnym zachodzie Europy (Kraj Basków, Bretonia, Anglia, Irlandia). Populacjom haplogrup I1, I2a2 i R1a1a można by przypisać dzieła kultury megalitów (kamiennych kręgów), w tym słynnego Stonehenge w Anglii z około 2800 lat przed Chr.
UWAGA! Obok Wenetów/Wenedów, stanowiących od starożytności wspólne etnosy z Germanami lub Słowianami, naukowa literatura wymienia także Wenetów iliryjskich na ziemiach północnej Italii (IX w. przed Chr.), później uległych romanizacji, oraz Wenetów Celtyckich, których etnonimy zostały zanotowane w Alpach Szwajcarskich, w Zachodniej Francji i w Walii. Dopatrzyć się ich można także pod nazwą „Enetoi” u Homera jako uczestników wojny trojańskiej w Azji Mniejszej przed 1200 r. przed Chr. i osadników nad Dolnym Padem we Włoszech, a później – w Eneidzie Wergiliusza (por. G. Jagodziński: Zagadkowy lud – Wenetowie; por. w Wikipedia)
| Populacje I1 i I2 to prawdopodobnie staroeuropejscy Wenetowie/Wenedowie (nie Wenecjanie!) Haplogrupa I wydzieliła się z ojcowskiej IJ może około 30.000-25.000 lat temu w Europie, zapewne na Bałkanach. Zapewne te populacje były twórcami przedlodowcowej kultury graweckiej i polodowcowej – magdaleńskiej. Przetrwały epokę wielkiego zlodowacenia zapewne w refugium nadwołżańskim (hg. I1) bałkańskim (hg I2a1), zachodniofrancuskim (hg I2b). Już w mezolicie i neolicie, rozproszone po całej Europie, łączyły się zapewne pod etnonimem Wenetowie/Wenedowie z napływowymi ludami, wyżej kulturowo zaawansowanych populacji R1a1a, a później także R1b1b2, tworząc z nimi nowe, krystalizujące się właśnie etnosy europejskie i wpływając niekiedy na kształt ich języków indoeuropejskich; najwyraźniej swój udział zaznaczyli w powstaniu etnosu germańskiego, dając mu zarazem znaczną część swojego zasobu językowego. Po kilku dziesiątkach tysięcy lat życia w rozproszeniu na kontynencie europejskim populacje I1 i I2 musiały bardzo różnić się językowo między sobą i jako lokalny substrat wpływały na różnicowanie się języków indoeuropejskich przybyłych tu populacji R1a (satemowych) i R1b (kentumowych). |
2. HAPLOGRUPA J1 i J2, pierwotnie bliskowschodnie.
Europejskie haplogrupy J1 i J2 są pochodzenia bliskowschodniego. Hg J1 w największym procencie występuje wśród Arabów, Żydów i Turków. Natomiast J2 jest dość powszechna w Europie południowej i południowo-wschodniej, zwłaszcza w rejonie Grecji (Turcja 33%, Kreta 32%, Albania 28%, Grecja 27%, Rumunia 24%, Italia 20%). Zapewne ona przybliżyła Europie elementy kultury fenickiej.
Głównie populacji podgrupy J2b2 (M241) zawdzięczamy przyniesienie rolnictwa z Anatolii do Europy na początku neolitu, około 7500 lat temu. Zaś do jego przejęcia i upowszechniania (m.in. w kulturze LBK) po Europie przyczyniły sie E1b1b1a2 i I2a2 (Battaglia et al. 2009) oraz na pewno populacje obydwu haplogrup R1a.
3. HAPLOGRUPA E1b1b1a, afrykańsko-fenicka, oraz E1b1b1a2 (południowoeuropejska).
Północno-wschodnia afrykańska populacja haplogrupy E, bardzo stara, bo powstała ok. 52.000 lat temu i żyjąca głównie w północno-wschodniej i północnej Afryce (zwł. Egipt i Etiopia), przybyła głównie przez małoazjatycką Anatolię do Grecji jako Fenicjanie, w okresie przedrolniczym, z Azji Mniejszej około 8000 lat przed Chr. Występuje zwłaszcza na południowych wybrzeżach Europy jako haplogrupa E1b1b1a (M78). Z niej, już na terenie Europy, wyłoniła się rdzennie europejska haplogrupa E1b1b1a2 (V-13). Populacje E znajdują się: Grecja 27%, Serbia 24%, Albania 22%, Portugalia 17%, Bułgaria 12%, Włochy 11%, Słowacja 11%. Zapewne ona przyniosła do Europy kulturę znad Nilu.
Haplogrupie E1b1b1a2 (V-13) przypisuje się, obok hg I2a2 (M423), przypisuje się upowszechnienie na Wybrzeżach Śródziemnomorskich rolnictwa, przejętego od przybyłej z Azji Mniejszej hg J2b2 (M241)
4. HAPLOGRYPA N1c i N1b, ugrofińskie.
Północnosyberyjskie i uralsko-fińskie populacje to haplogrupa N (na drzewie genealogicznym hg N to jakby daleka kuzynka haplogrupy R). Haplogrupa N wyłoniła się we wschodnich Chinach około 12.000 lat temu, a jej podgrupy przez rejon Ałtaju dotarły do ziem Syberii podbiegunowej, a stąd około 6-7 tysięcy lat temu (AK) do północnowschodniej Europy. Licznie reprezentowana podgrupa N1c (M46/Tat) datowana jest na około 1500 lat przed Chr., głównie w północnowschodnich rejonach Europy od Uralu i górnej Wołgi po północną Skandynawię (Suomi-Lapończycy). I tak: Finlandia 58%, Litwa 42%, Łotwa 38%, Estonia 34%, Rosja 23%, a wśród Węgrów – tylko 1% (co ujawnia, że w zdobyciu ziem słowiańskich nad Dunajem w końcu IX w. uczestniczył tylko niewielki oddział rdzennych Madziarów). Daleko mniej liczna występuje wśród nich także siostrzana haplogrupa N1b (M43).
5. HAPLOGRUPA G2a, pierwotnie kaukaska.
Sporadycznie i w niewielkiej liczbie występuje w Europie ludność haplogrupy G2a. Przybyła ona z Azji Mniejszej, głównie może z populacji Ormian (tam około 30-40%) i sąsiednich populacji oraz Turcji (kilkanaście procent). W znacznym procencie zaludniają dziś Sardynię (15%). Poniżej 10% zaludniają Szwajcarię, Austrię, południowe Niemcy i Czechy. Nie jest wyjaśniona obecność i rola tej populacji w Europie. Prawdopodobnie upowszechniali oni wyroby z miedzi w Europie w okresie wczesnego neolitu i przed epoką brązu, czyli około 4000 lat przed Chr.
6. HAPLOGRUPA H1a, hindusko-cygańska.
Kilka innych pozostałych nieindoeuropejskich haplogrup w Europie, bardzo rzadkich, ma tu pochodzenie raczej współczesne. Najliczniejsza z nich to głównie pochodząca z Indii (Pendżab) haplogrupa H, którą odznaczają się europejscy Cyganie i pokrewne im plemiona (np. Romowie). Ich przybycie do Europy datowane jest na około IX wiek. Ich język należy jednak do grupy indoirańskiej z półwyspu Indyjskiego.
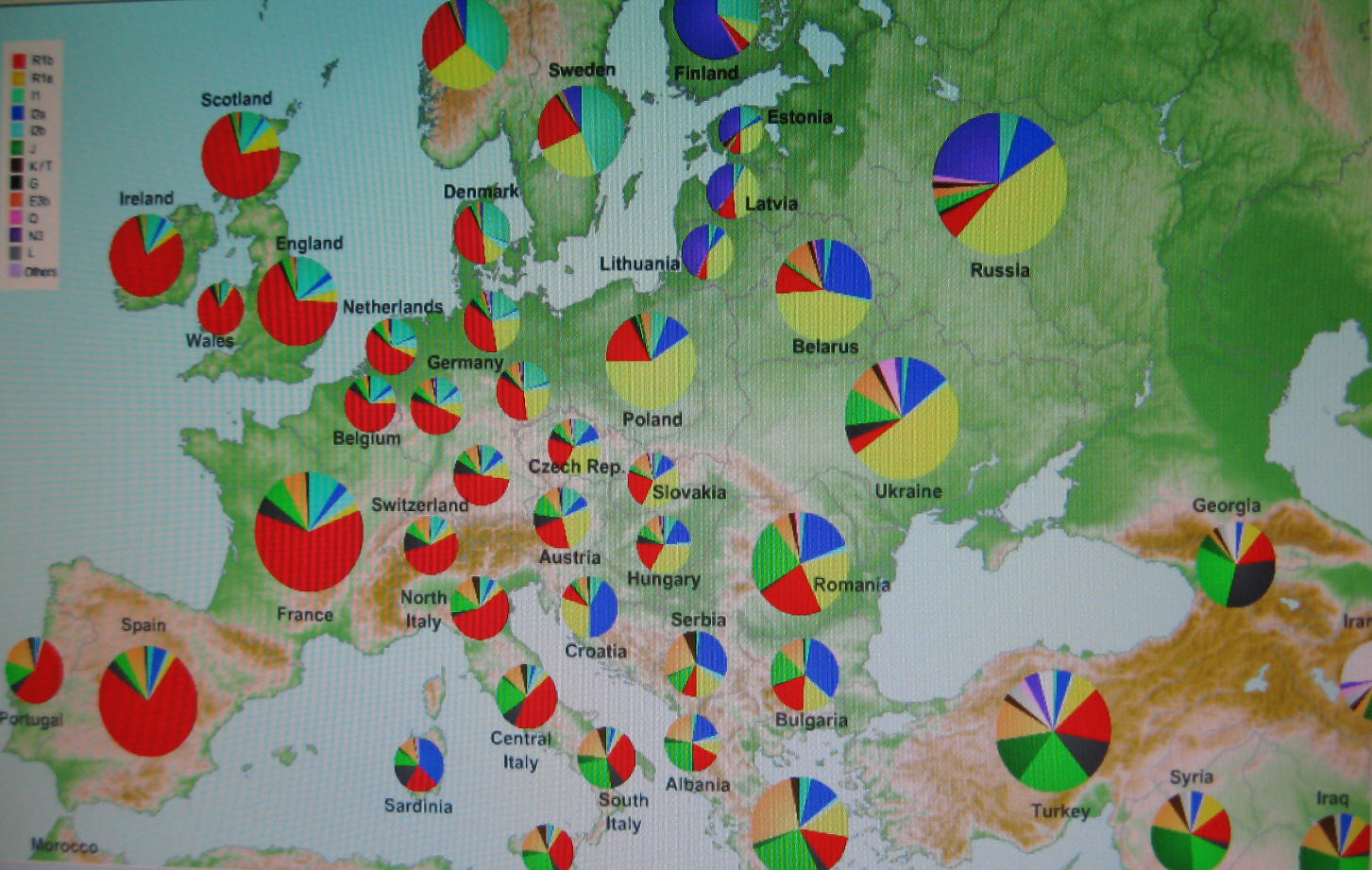 |
Dzisiejsze rozmieszczenie męskiego Y-DNA w Europie (według portalu Eupedia.tabl) Kolor żółty R1a1a (wschodnioeuropejski, słowiański); czerwony R1b1b12 (zachodnioeuropejski, celtycko-germański); modry I1 (nordycko-germański); granatowy I2a (bałkańsko-słowiański); błękitny I2b (bałkańsko-niemiecki), pomarańczowy E (północno-wschodnioafrykański i grecki); fioletowy N (uralsko-fiński, północnorosyjski), zielony J1 (semicki) i J2 (bliskowschodni), czarny G (kaukaski). |
Rys. 6
Powyższe haplogrupy, choć genetycznie nieindoeuropejskie, w znacznej mierze przyczyniły się nie tylko do powstania dzisiejszego kształtu języków indoeuropejskich w Europie, ale także całego bogactwa kultury duchowej i materialnej Europy, każda z nich w swoim zakresie i według własnych możliwości populacyjnych i czasowych.
MITOCHONDRIALNE DNA
WSKAZUJe MATCZYNE POCHODZENIE CZŁOWIEKA
DNA mitochondrialne, czyli mtDNA, to materiał genetyczny znajdujący się w rejonie pozajądrowym komórek. Białka kodowane przez mtDNA to część białek łańcucha oddechowego (regulują energetykę komórki i całego organizmu).
DNA mitochondrialne są wykorzystywane w genealogii genetycznej i medycynie sądowej. Służą do tego niekodujące fragmenty (regiony) genomu mitochondrialnego o wysokim zróżnicowaniu i zmienności u poszczególnych ludzi.
Szybkozmienny region HVR1 obejmuje sekwencję 16001-16568, a region HVR2 – sekwencję 001-574.
Porównanie DNA mitochondriów pochodzących od ludzi, wywodzących się z różnych grup etnicznych, pozwoliło na obliczenie, kiedy żyła tzw. Ewa mitochondrialna – kobieta, od której wywodzą się wszyscy współcześni ludzie, a właściwie ich genom mitochondrialny. Oto niżej (z Wikipedii) mapa mutacji i migracji haplogrup mtDNA od mitochondrialnej Ewy z Afryki na resztę świata.
Powstała na Bliskim Wschodzie haplogrupa mtN i liczne jej podgrupy znajdują się, w przeciwieństwie do haplogrupy mtM i jej podgrup, głównie w rejonie Kaukazu, Azji Mniejszej i w Europie; zapewne one uczestniczyły w powstaniu ludzkiej rasy kaukaskiej, europidalnej, białej.

Natomiast haplogrupa mtM i jej podgrupy – upowszechniły na pozostałych kontynentach (poza Afryką).
Na poniższej, ogólnoświatowej mapie brak wytworzonej z haplogrupy N i w jej regionie, a niedawno odkrytej, haplogrup mtR (zob. wyżej!), od której wywodzą się prawie wszystkie podgrupy europejskie, a zwłaszcza poprzez mtRO i mtHV ważna europejska haplogrupa H. Obejmuje ona w ponad 20 podgrupach około 40% ludności europejskiej i polskiej. O obecności haplogrup mitochondrialnych w rejonie Dunajca, zob.
W moim rodzie podgrupą mtHV-1 wyróżniała się Katarzyna Kuras Szlagowa z Łukowicy, popularna w Europie podgrupę H miała Anastazja Wilk Gurgulowa ze Stronia, a podgrupą mtH-14a odznaczała się Katarzyna Iwan Szlagowa z Łukowicy.
Z haplogrupy mtR pochodzi też podgrupa mtU, bardzo stara w Europie. Grupą U3 wyróżniała się w moim rodzie Anna Legutko Janikowa, rodem z Jasiennej i Łazów Biegonickich.
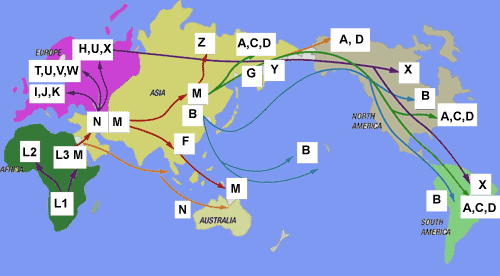
MtDNA przekazywane jest zarówno synom, jak i córkom, ale tylko przez matki. Dlatego służą do rozpoznania tylko matczynego pochodzenia, zarówno mężczyzn, jak i kobiet; podczas gdy Y-DNA służy poznaniu tylko ojcowskiego pochodzenia, i tylko mężczyzn. MtDNA ułatwiają też rozpoznanie tras dawnych migracji rodów i plemion, choć nie tak wyraziście, jak męskie Y-DNA; mężczyźni bowiem ściślej tkwili w swoich pierwotnych grupach rodowych i plemiennych.
Np. mtDNA-H5 i H7, stwierdzane w rejonie Kaukazu, wzdłuż Dunaju i pod Alpami, migrowało z męską Y-hg. R1b. Podobnie mt-U3, stwierdzane wokół Morza Czarnego oraz mt-J, rozpowszechnione pierwotnie w Azji Środkowej i wokół mórz Kaspijskiego i Czarnego. Haplogrupie Y-R1a towarzyszyło głównie mt-U4, bardziej popularne w rejonie Tadżykistanu i Pakistanu, w Azji Środkowej oraz w Europie Wschodniej. Pochodząca z Afryki północno-wschodniej mt-T, stwierdzana jest w rejonie Morza Egejskiego i Europie Wschodniej; z populacjami Y-R1a można łączyć też mt-U3 i mt-J (towarzyszące jednak głownie Y-hg. R1b). Mt-H13 (i mniej H2) łączy Kaukaz i Sardynię, podobnie jak męska Y-G2a. Mt-H1, H3, V i U5 zdradzają paleolityczne i mezolityczne towarzystwo łowców-zbieraczy (Y-hg. I); także mt-J1 występuje przy germańskich populacjach hg. I. Mt-K1a towarzyszy głównie męskim J, T i E1b1b.
Ponieważ mitochondrialne DNA łatwiej niż Y-DNA trwają niezniszczalnie w szczątkach kopalnych, mogą ułatwić identyfikację oraz stwierdzenie ciągłości między człowiekiem paleolitycznym a dzisiejszym, np. człowiek z Kostienek (nad Donem w Rosji), z około 30.000 lat temu, odznaczał się haplogrupą U2; mtDNA kromaniońskiego człowieka z Paglicci Cave (Włochy), z około 28.000 lat temu, ujawniło jednoznaczną różnicę względem neandertalczyka, a tożsamość z dzisiejszymi typami HV lub U.
Genetycy starają się też ustalić haplogrupy, wyróżniające populacje słowiańskie w Europie; zob. np.:
1) Grzybowski et al. 2007: Complex interactions of the Eastern and Western Slavic populations with other European groups as revealed by mitochondrial DNA analysis.
2) Malyarchuk et al. 2008: Mitochondrial DNA Phylogeny in Eastern and Western Slavs.
3) Malyarchuk et al. 2010: The Peopling of Europe from the Mitochondrial Haplogroup U5 Perspective
Ilustracją obecności poszczególnych haplogrup mtDNA na polskiej ziemi są rezultaty analiz w ramach polskiego programu/projektu Family Tree DNA, który administruje L. Mayka (tablica może otwierać się powoli, podobnie jak tam dołączona mapa).
 |
Polskie mtDNA w testach FTDNA |
W sumie, inaczej niż ojcowski rodowód, matczyny rodowód Polek i Polaków przeważnie nie różni się od pozostałych krajów Europy (zob. na tablicy Eupedii). Wyraźne różnice zachodzą natomiast w stosunku do ludności krajów Azji i Afryki, jak wskazuje mapa migracji.
Zob. najnowsze opracowanie datowania mitochondrialnych haplogrup: Soares et al. 2009.
Zob. tenże, tablica datowania, Supplemental data: An Improved Human Mitochondrial Molecular Clocka.
Zob. praktyczne zastosowanie genealogii mtDNA na rodzinnej stronie: Szlagowie.
a) Dlaczego tak szczegółowo, jak na internetowe zwyczaje, z podaniem podstawowej terminologii i literatury, przedstawiam ten temat?
Dlatego, że dla mnie i wielu z nas bardzo ważne jest pytanie: Jaka jest nasza genealogia – skąd jesteśmy, co nas łączy lub dzieli, jaka jest nasza przeszłość, dlaczego nie możemy się jej wyrzekać, a wreszcie dlaczego nie powinniśmy pogardzać swoją słowiańskością i polskością!
Genealogia Y-chromosomowa (Y-DNA) bardzo służy historii, poznaniu genezy pierwotnych ludzkich plemion, większych populacji i narodów oraz ich przemieszczaniu się po Ziemi. Ułatwia poznanie wydzielonych męskich rodów i w ogóle rodzin, które nas wydały na świat, do których należymy i wobec których mamy obowiązki – nie tylko wdzięczności! Zdecydowanie pomaga określić twórców archeologicznych kultur, ujawnianych w pracach wykopaliskowych.
Ma to szczególne znaczenie prestiżowe dla Słowian. Rzekome późne wyłonienie się etnosu słowiańskiego (przypominam: gr. „etnos” to rój, stado, plemię, a nie sama kultura!) bywało nieprzychylnie dla Słowian interpretowane. Do tego przyczyniło sie także i nazewnictwo. Mianowicie, ponieważ we wczesnym średniowieczu Słowianie bywali przez kupców arabskich i żydowskich sprzedawani na rynkach Europy jako niewolnicy, od etnonimu Słowianin, łac. Sclavus, utworzono około X w. łacińskie słowo „sclavus”, oznaczające niewolnika, sługę, a także w ogóle człowieka gnuśnego. I to określenie w rozmaitej formie (np. sclave slave, esclave, slav…) przedostało się do innych języków zachodnich. Niekiedy niestety wpływa ono na opinię o ludach słowiańskich i na sposób traktowania Słowian przez zachodnich Europejczyków.
b) Rozpoznawanie swojego cennego rodowodu w ojcowskim Y-DNA czy macierzyńskim mtDNA i wydzielanie krewniaczych haplogrup ludzkich nie oznacza tworzenia jakiegoś nowego rasizmu, tym razem genetycznego, na miejsce dotychczasowego, opartego zasadniczo tylko na zewnętrznym wyglądzie człowieka. Genealogia Y-DNA czy mt-DNA umożliwia zidentyfikowanie i umieszczenie osób czy rodu na ojcowskim czy macierzyńskim drzewie genealogicznym ludzkości i ustalenie czasu wyodrębnienia się, co w sumie już daje wielkie poczucie zakorzenia i wspólnoty, ale to dopiero ułamek prawdy o pochodzeniu konkretnej osoby.
Bowiem oprócz niewielkiego chromosomu płciowego Y-DNA lub mitochondrialnego mtDNA posiadamy daleko większe 22 inne pary chromosomów, tzw. autosomalnych, i ponad trzydzieści tysięcy innych genów kodujących. Przekazywane są one zarówno przez ojca, jak i przez matkę, i zarówno synom, jak i córkom. One kształtują podstawowe funkcje organizmu, a także kolor skóry, oczu, czy inne nawet bardzo drugorzędne drobiazgi. W ten sposób każdy dzisiejszy człowiek ma pozostałe chromosomy i geny nie tylko od około 8.000 swoich dotychczasowych przodków w linii prostej aż do Adama i Ewy, ale także, jak obliczyli antropologowie, od około 80.000 innych przodków z bardzo rozbudowanych linii bocznych.
Świadomość tego, że nosimy w sobie materiał genetyczny tylu, z rozmaitych haplogrup i ras przodków, powinna – niezależnie od ustalonej haplogrupy Y-DNA lub mt-DNA – chronić od rozmaitych ideologii rasistowskich i pozytywnie wpływać na poczucie braterstwa z wszystkimi istotami ludzkimi. Genetyka antropologiczna już poszukuje w autosomach, czyli chromosomach pozapłciowych, owych mutacji „rasowych”, czyli kształtujących odrębny wygląd i cechy fizyczne poszczególnych grup ludzkich oraz ustala na mapach świata trasy „migracji” dziedziczonych genów, tak jak już od dawna czyni to genetyka medyczna, identyfikująca obciążone chorobami dziedzicznymi osoby i populacje.
c) Ponieważ wraz z odkrywaniem nowych mutacji w SNP zmienia się klasyfikacja haplogrup na drzewie genealogicznym, jak ostatnio zmieniono R1a1 na R1a1a i R1a1a7 i inne, a nadto stale napływają nowe etniczne i geograficzne ustalenia co do poszczególnych populacji, dlatego co pewien czas może zajść potrzeba dokonania poprawek w tym artykule; mogą też powstać niezgodności z poprzednimi tekstami i cytowanymi pracami. Trzeba też śledzić zmiany w terminologii, nazewnictwa i symboli haplogrup. Np. dla uproszczenia umownie podaje się krótszą nazwę haplogrupy ojcowskiej zamiast wszystkich synowskich, np. ojcowską R1b zamiast kilkudziesięciu haplogrup potomnych, ze skomplikowanymi ich określeniami; albo R1b1b2 dla określenia wszystkich europejskich jej podgrup. Niekiedy podaje się tylko krótsze określenie mutacji, zamiast dłuższego określenia odpowiedniej haplogrupy – miejsca na drzewie genealogicznym, np. R-M458 zamiast R1a1a7 albo R-U152 zamiast R1b1b2a1a2d – alpejskich i italskich Celtów. Nadto w literaturze, niekiedy niekonsekwentnie, dodaje się gwiazdkę, np. R1a*, dla określenia, że nie bierze się pod uwagę jej podgrup.
Uzupełnienie 08 08 2016
AKTUALNE PRACE GENEALOGII GENETYCZNEJ
L. Mayka prowadzi program FTDNA Polish Project . Rejestruje tam się wyniki analiz przeprowadzonych w pracowniach FamilyTreeDNA dotyczących materiałów z DNA, dostarczonych przez Polaków; internetowa strona przedstawia rezultaty badań i identyfikacje haplogrup Polaków, a w haplogrupie R1a1a1g – podział na klastry, w zależności od haplotypów STR.
Dokładnym rozpracowaniem polskich haplotypów z haplogrup R1a1a i R1a1a1g zajmuje się P. Gwozdz. Na swojej internetowej stronie Polish clades prowadzi analizę haplotypów, grupuje, oblicza czas populacji i ich migracji. W opublikowanej w JOGG Journal of Gnetic Genealog 5/2 2009 jego pracy i pojawia się więcej informacji na temat metody pracy i jej rezultatów (część II). Czas wspólnego przodka haplotypu P .
Prace zespołu Łukasza Łapińskiego.
Całością haplogrupy R1a jej mutacji i podgrup w świecie oraz podziałem na genetyczne gałęzie zajmuje się od czerwca 2011 r. genealog Łukasz Łapiński na stronie R1a and Subclades Y-DNA Project.
(Natomiast podziałem tego rodu na poszczególne kraje lub większe grupy narodowościowe i regionalne zajmuje się strona Cyndi (Kanada): R1a Y-chromosome Haplogroup Project.)
|
GENEZA JĘZYKÓW SATEMOWYCH (hindo-bałto-słowiańskich) Na ojczystym terenie, zapewne gdzieś w rejonie Dunaju lub Karpat, w populacji R1a1a1 dokonała się satemizacja języka (dotąd zapewne kentumowego jak nadal w haplogrupie R1b), równoznaczna z formowaniem się języka praindo-prabałto-prasłowiańskiego. Satemizacja polegała głównie na innowacji, że pierwotnie miękkie k’ u satemowców przeszło w s (u kentumowców tylko utraciło zmiękczenie), stając sie zwykłym k lub c. Wnet potem, od około III tysiąclecia p.n.e. (ale jeszcze przed terytorialną ekspansją Prasłowian), nastąpiło oddzielenie się języka indo-irańskich Ariów (głównie z mutacją Z93), a później oddzielenie się języka bałtyjskiego. |
UWAGA.
W rejonie Europy Środkowej nastąpiło spotkanie ludności hg. R1a1a z ludnością kultury pucharów lejkowatych (zapewne hg. I1 i I2), oraz zainicjowanie i/lub upowszechnianie kilku innych kultur, zwłaszcza kultury amfor kulistych i ceramiki sznurowej (Eulau) z toporami gładzonymi i grobami jednostkowymi. Regres demograficzny, trwający w drugim tysiącleciu przed Chr. w rejonie Europy Środkowej, nie był całkowity i nie oznaczał katastrofy kulturowej, zwłaszcza że na sąsiednich ziemiach dzisiejszej Ukrainy i Rosji Europejskiej tego regresu nie było, co umożliwiło późniejsze wsparcie demograficzne i kulturowe dla ziem rejonu Środkowej Europy.
Postscriptum.
Powyższe badania zdają się nie potwierdzać tezy niektórych archeologów o naddnieprzańskim pochodzeniu Polaków i ich haplogrupy R1a, i to dopiero w V wieku po Chr. Co więcej, wyraźnie zauważa się, że Polska leży jakby w centrum starożytnego osadnictwa słowiańskiego haplogrupy R1a. W Rosji np. o rezultatach pobierania próbek DNA wprost pisze się: Poskrob Ruskiego – znajdziesz Polaka ( Поскреби русского – найдешь поляка, Газета.ru 14.01.2008) – jak przedtem, podobno niesłusznie, tam się mawiało: Поскребешь русского – найдешь татарина – 'Podrapiesz Ruskiego, znajdziesz Tatara’.
* * * * *
Archeologiczne (ancient) Y-DNA w Europie.
Tabela i ilustracja zasiedlenia starożytnej Europy według znalezisk archeologicznych Y-DNA;
w kulturach, czasie i procentach (stan badań grudzień 2015)
(źródło: Eupedia, Tomenable/Davidski)
http://s14.postimg.org/mu6jakwv5/Chronological_YDNA.png
{kind=link}
http://www.tropie.tarnow.opoka.org.pl/images/ancient.ydna.eur.png
(klik-powiększenie)
Dyskusja
-
B. czyzowa said, on 21 Marzec 2011 at 17:41 (Edytuj)
Mili Państwo! Mam znajomą w Moskwie, która bardzo interesuje się m.in. genealogią Słowian, ma ogromny szacunek dla Polaków, którzy – wg jej rozeznania – zachowali w nazewnictwie bardzo dużo nazw starosłowiańskich (w odróżnieniu m.in.. od Rosjan). Chciałaby nawiązać kontakt z kimś, kto zna tę tematykę. Korespondować może jednak tylko po rosyjsku. Na wszelki wypadek podaję jej adres mailowy: korobin@rambler.ru. Ma na imię Natalia.
Pozdrawiam i
B. CzyżowaOdpowiedz-
bialczynski said, on 21 Marzec 2011 at 17:49 (Edytuj)
Postaramy się nawiązać kontakt, dziękujemy. Pozdrawiamy.
C.B.
Odpowiedz
-
-
krt said, on 21 Lipiec 2011 at 20:14 (Edytuj)
Z zainteresowaniem czytam stronę i chcę podzielić się kilkoma uwagami, które może podsuną autorowi pomysły do nowych teorii.
1 sprawa.
W średniowieczu polscy kronikarze wywodzili pochodzenie lechitów z Panonii. Tereny obecnej Austrii. Brzmi na pierwszy rzut oka niedorzecznie, ale:
Na tych terenach jest rzeka Lech !!!!
Lech (łac. Licus, Licca) – rzeka w Austrii i Niemczech, prawy dopływ Dunaju. Długość – 264 km, powierzchnia zlewni – 4.126 km².
Góra Zermatten (Sarmaci).I patrzmy wyżej rzeka Lech to po łacinie Licus, Licca, czy to nie nasuwa skojarzeń z Lechitami – Licikaviki ?
Licikaviki – nazwa występująca tylko jeden raz w Dziejach saskich Widukinda z Korbei. Autor kroniki wspomina, że było to plemię słowiańskie podlegające władzy Mieszka I. Pisze o nim przy okazji relacjonowania ataków Wichmana na państwo Polan.
2 sprawa
Dagome iudex
Tłumaczenie polskie tekstu:
Podobnie w innym tomie z czasów papieża Jana XV Dagome, pan, i Ote, pani i synowie ich Mieszko i Lambert (nie wiem, jakiego to plemienia ludzie, sądzę jednak, że to byli Sardyńczycy, ponieważ ci są rządzeni przez czterech “panów”) mieli nadać świętemu Piotrowi w całości jedno państwo, które zwie się Schinesghe z wszystkimi swymi przynależnościami w tych granicach, jak się zaczyna od pierwszego boku wzdłuż morza [dalej] granicą Prus aż do miejsca, które nazywa się Ruś, a granicą Rusi [dalej] ciągnąc aż do Krakowa i od tego Krakowa aż do rzeki Odry, prosto do miejsca, które nazywa się Alemure, a od tej Alemury aż do ziemi Milczan i od granicy Milczan prosto do Odry i stąd idąc wzdłuż rzeki Odry aż do rzeczonego państwa Schinesghe.[4]
Może to nie chodzi o Mieszka I i Gniezno, nie o jakiegoś dziwnego dagome iudex, a o degomir dux, czyli i Tęgomir, np. ten:
Tęgomir (zm. po 938) – książę Stodoran.
Jesienią 928 na ziemie Tęgomira najechał król niemiecki Henryk I Ptasznik. Książę wycofał się do Brenny, gdzie został oblężony przez wojska Henryka. Walki pod słowiańskim grodem zakończyły się klęską Tęgomira. Pokonany książę został uwięziony, zaś plemię Stodoran zmuszone do opłacania trybutu. Będąc w niewoli, Tęgomir zaprzyjaźnił się z dawnymi wrogami, zaś jego córka została prawdopodobnie nałożnicą syna Henryka, Ottona[1]. Ze związku tego miał pochodzić nieślubny syn, arcybiskup Moguncji Wilhelm. Po wyjściu z niewoli niemieckiej w 938 Tęgomir wziął udział w zmowie z margrabią Marchii Wschodniej Geronem. Przybył do Brenny, gdzie ogłosił swój powrót. Współplemieńcy uznali jego władzę i przywrócili mu tron książęcy. Wkrótce Tęgomir podstępnie zgładził swego nieznanego z imienia bratanka, jedynego dziedzica tronu stodorańskiego, i poddał się wraz z ludem władzy Ottona.
No skoro poddał się władzy Ottona to i dla równowagi mógł się poddać papieżowi, żeby pokazać że przyjął chrześcijaństwo.
3 sprawa
Czeska legenda o Kroku i polska o Kraku – dotyczą wg mnie tej samej osoby, wynika z tego że Słowianie szli z zachodu na wschód (znowu wraca teoria o Panonii, z rzeka Lech, górą Zermaten, itd.) i część została w Bohemii, część w Polsce , reszta poszła dalej na wschód przez Rosję aż do Indii, w Pakistanie tez sa nazwy Leh i inne znajomo brzmiące).
Przy takim założeniu nabiera kolorów wersja o słowiańskich Etruskach i ich runach, w końcu z Panonii do Płw. Apenińskiego niedaleko.
Co do nazwy Krak, Kraków, to w Chorwacji aż się roi od nazw Krk, Krkova Hora itp.
Przy czym znamienne jest że w wersji czeskiej jest tylko Czech i Lech, a polskiej i ruskiej dochodzi Rus. Czyli że rozdzielanie się następowało stopniowo.
4 sprawa
Nazwa “Polska” może to wpływ wikingów, tak jak “Skanska”. Czy to nie dziwne, że Mieszko I wydaje córkę za wikinga?
Sygryda Storråda, Świętosława, Saum-Aesa (szw. Sigrid Storråda, w Danii jako Gunhild czyli Gunhilda, po słowiańsku Świętosława) (ur. między 960 – 972, zm. po 1016) – królowa Szwecji, Danii i Norwegii. Córka Mieszka I i prawdopodobnie Dobrawy, siostra Bolesława Chrobrego, w sagach skandynawskich wzmiankowanego jako “Burisleif lub Burislafr, potężny konung Windlandii”.
Kilka średniowiecznych kronik stoi na stanowisku, że matka Haralda II i Kanuta Wielkiego wywodziła się z plemienia Polan lub innego blisko z nimi spokrewnionego plemienia słowiańskiego. Ten rodowód Sygrydy potwierdzają:* Thietmar z Merseburga wspomina, że córka Mieszka I, a siostra Bolesława Chrobrego wyszła z mąż za Swena Widłobrodego i urodziła mu dwóch synów, Haralda II i Kanuta Wielkiego. W przekazie tym nie ma jednak żadnej wzmianki na temat jej imienia. Thietmar miał prawdopodobnie największą wiedzę spośród wszystkich średniowiecznych kronikarzy na temat wydarzeń mu współczesnych, ponadto był dosyć dobrze zaznajomiony z sytuacją w Polsce i Danii w tamtym czasie.
* Adam z Bremy pisze, że polska księżniczka była żoną Eryka Zwycięskiego i matką Haralda II oraz Kanuta Wielkiego. Informacja ta jest uważana przez niektórych historyków za niepewną.
* Encomium Emmae Reginae zawiera wzmiankę, że Kanut Wielki i jego brat przybyli na ziemie Słowian po swoją matkę, żeby zabrać ją z powrotem do Danii. Informacja ta nie przesądza tego, że Sygryda wywodziła się z plemion słowiańskich, jednak kronika mocno to sugeruje.
* Liber vitae of the New Minster and Hyde Abbey Winchester zawiera zapis mówiący, że siostra króla Kanuta nosiła imię Santslaue (Santslaue soror CNVTI regis nostri), które bez wątpienia jest imieniem słowiańskim. J. Steenstrup sugeruje, że siostra Kanuta została tak nazwana po swojej matce, stąd hipoteza (odtąd uważana za słuszną), że słowiańską wersją imienia Sygryda jest Świętosława. To stwierdzenie pokrywa się z tezą, jakoby Sygryda byłą córką Mieszka.Twierdzenie, że matką Kanuta była siostra Bolesława Chrobrego wyjaśnia kilka zagadkowych zapisów w średniowiecznych kronikach, jak choćby to, że polscy wojownicy brali udział w podboju Anglii.
Później kontakty z północą się nagle kończą. Czy to nie zastanawiające?
Historia Słowian jest niezwykle ciekawa, aż dziw jak Polacy mało o niej wiedzą.
Gratuluje artykułów o runach słowiańskich, ciekawe kiedy dotrze ta wiedza do szerszego audytorium
Odpowiedz-
Dziękuję za bardzo inspirujący i wnikliwy wpis. W Księdze Ruty umieściłem Nową Kolibę Kruków- Słowian czyli Lęgię Naddunajską w takiej a nie innej lokalizacji bo ufam kronikarzom, a nie ufam tym którzy dla popisów erudycyjnych gotowi są zakwestionować rozum Starożytnych. Kiedy pisałem Księgę Tura i Księgę Ruty Internet zalewały komentarze typu że Białczyński to fantasta, który sobie wymyśla bajeczki. Dzisiaj genetyka bezspornie potwierdza moje dedukcje i intuicje. To pokazuje, że trzeba więcej pokory w studiach nad przeszłością i że prywatne studia interdyscyplinarne prowadzone przez wiele lat pozwalają wciąż odkrywać na nowo świat – odkrywać go z sensem i pożytkiem dla Prawdy Historycznej, a PRZECIW HISTORYCZNEJ PROPAGANDZIE.
Cały czas pamiętajmy że naszą historię piszą od 1000 lat najeźdźcy bo Lęgia-Lechia – Polęgia – Polechia- Polsza-Polska jest krajem frontowym, a przez ostatnie 250 lat historię Słowian pisali Niemcy, a historię Polski Rosjanie. A był to okres bezprecedensowego rozwoju nauki, narzędzi naukowych i cywilizacji europejskiej. Wszystko to było podporządkowane propagandzie najeźdźców. Mało jest krajów na świecie które były pod kolonialnym uciskiem przez 250 lat.
Pozdrawiam – Pańskie spostrzeżenia powinny się tu znaleźć w formie artykułu, bo są bardzo ważne merytorycznie.
CB
-