
GENETYCY NA TROPIE POCHODZENIA EUROPEJCZYKÓW, CZĘŚĆ 1
WSTĘP
W bieżącym 2017 r. ukazało się wiele prac genetyków opisujących historię pochodzenia Europejczyków. Prace te zawierały liczne próbki kopalnego DNA, czyli pobrane ze szkieletów ludzi zamieszkujących nasz kontynent tysiące lat temu. Temat opublikowanych prac naukowych jest bardzo szeroki. Analiza danych w nich zawartych mogłaby zająć setki stron. Ponadto wciąż powstają kolejne prace naukowe przez co wciąż poszerza się pole dyskusji.
Niniejszy artykuł nie jest ani streszczeniem ani recenzją powstałych prac naukowych. Komentowanie tekstów naukowych jest bowiem niekończącym się zajęciem. Miejscem takich komentarzy są m.in. fora naukowe. Dlatego w artykule skupiam się na tym, co z punktu widzenia odkrywania prawdy o pochodzeniu Europejczyków jest według mnie najważniejsze. Mam tu na myśli analizę próbek genetycznych, a konkretnie ich haplogrup, subkladów i mutacji. Ponieważ w tym celu najbardziej wyrazisty obraz daje analiza próbek męskiego chromosomu Y (tzw. Y-DNA), to swoją krótką analizę opieram właśnie na nich. Dotyczy to także próbek pochodzących z Polski, których niestety wciąż jest bardzo mało, co utrudnia, ale nie wyklucza dotarcia do prawdy o pochodzeniu Polaków. Mimo, iż wciąż brakuje kompleksowego opracowania dotyczącego historii genetycznej dawnych mieszkańców Polski, to jednak pewne wnioski nasuwają się z dotychczasowych badań.
Temat badań genetycznych jest zagadnieniem bardzo szerokim i stale rozwijającym się. Artykułów na ten temat można by napisać nieskończoną ilość. Wziąć też należy pod uwagę fakt, iż następne badania dopiero są prowadzone, a jeszcze inne powstaną w przyszłości. Mając tego świadomość, pozostawiam ten temat otwarty. Stąd tytuł artykułu opatrzony jest wyróżnikiem „część 1”. Kolejne części tego zagadnienia mogą bowiem powstać. W najbliższym czasie planuję opublikowanie kolejnego artykułu związanego z badaniami genetycznymi i odkrywaniem pochodzenia mieszkańców Europy. Tytuł tego kolejnego artykułu będzie taki sam jak niniejszego, z tą różnicą, że będzie on uzupełniony dopiskiem „część druga”. Jest kwestią otwartą czy powstaną kolejne części tej „genetycznej” serii. Tematów z dziedziny genetyki populacyjnej na pewno nie będzie brakować do omówienia. Wszystko zależeć będzie od woli i determinacji w pisaniu kolejnych części. W każdym razie pozostawiam sobie otwartą furtkę, że w każdej chwili kolejne artykuły o tym zagadnieniu mogą powstać.
Część pierwsza, niniejsza, skupia się na czterech rozprawach międzynarodowych zespołów naukowych. Poniżej przedstawiam ważne teksty oparte o badania genetyczne, którymi przewodzili: Iain Mathieson, Mark Lipson, Alissa Mittnik i Lehti Saag. Przy okazji zahaczam o kilka innych genetycznych prac naukowych, powstałych we wcześniejszych latach.
IAIN MATHIESON Z ZESPOŁEM, 2017 r.
Opracowanie zaczynam od jednej z najważniejszych prac, jakie ukazały się w bieżącym 2017 r. Mowa o pracy zespołu pod przewodnictwem Iaina Mathiesona, genetyka z Harvard Medical School w Bostonie. [1] Rozprawa, o której mowa zawiera niebywałą wręcz ilość próbek kopalnego DNA. Jest ich ponad 200, z czego 129 to próbki męskiego DNA. Próbki pochodzą z 12 państw, w tym 6 pochodzi z Polski. Jedna próbka jest spoza Europy – z tureckiej Anatolii.
FRANCJA:
Najstarsze próbki wyodrębnione przez zespół Mathiesona pochodzą z Francji. Niestety ich jakość pozostawia wiele do życzenia, gdyż wyniki są mało precyzyjne, a nawet w jednym przypadku – niepewne. Są to trzy próbki o haplogrupach: I, J? (próbka niepewna) oraz R (bez skonkretyzowania czy jest to R1a czy R1b, a nawet czy w ogóle chodzi o R1). Datowane są one na lata: 11.140 – 9460 p.n.e. Oprócz tych trzech próbek, na terenie Francji wyodrębniono czwartą – datowaną na lata 5370 – 5220 p.n.e. Jest to próbka o haplogrupie I. Wszystkie cztery francuskie próbki związane są z ludnością łowców-zbieraczy.
NIEMCY, AUSTRIA, POLSKA:
Lepsze wyniki zespół amerykańskiego genetyka uzyskał w przypadku próbek z obszaru położonego na wschód od Francji, czyli z Europy Centralnej – Niemiec, Austrii i Polski. Najstarsza próbka z tej części Europy pochodzi z okresu 7460 – 7040 lat p.n.e. Wydobyto ją ze szkieletu spoczywającego w jaskini Falkenstein (niem. Falkenstein-Höhle) w Badenii-Wirtembergii (Niemcy). Jej Y-DNA to I2a2a (zachodni krąg łowców-zbieraczy).
Kolejne 6 próbek pochodzi głównie z szóstego tysiąclecia (dokładnie z przedziału lat 5500 – 4500 p.n.e.) z Austrii i Niemiec. Oto ich haplogrupy: jedna J2, jedna C1a2 oraz cztery G2a (2 x G2a2a, 1 x G2a2a1, 1 x G2a2b2a3).
Następne w chronologii to 6 próbek z Polski, wszystkie z miejscowości Kierzkowo (woj. kujawsko-pomorskie, pow. żniński). Datowane są głównie na czwarte tysiąclecie, a dokładnie na lata 3400 – 2575 p.n.e. Ludność, od której pobrano te próbki związana była z kulturą amfor kulistych. Polski archeolog, prof. Witold Hensel uważał Kujawy za kolebkę tej kultury. [2] Cztery spośród sześciu próbek związane są z haplogrupą I2 (1 x I2, 2 x I2a2, 1 x I2a2a1b), jedna z nich to hg CT i jedna BT. Oznacza to, że wspomniana kultura związana była z ludnością mezolityczną Europy (haplogrupy: I, C, H, T). Jak na razie nie znaleziono na tym stanowisku ludności wiązanej z językami indoeuropejskimi (hg R1a i R1b).
Dopiero najmłodsza próbka opisana przez zespół Mathiesona to próbka o haplogupie R1a (R1a1a1b1a2) pochodząca z Halberstadt-Sonntagsfeld w Niemczech, datowana na lata 1193 – 979 p.n.e. Próbka ta oznaczona jest numerem identyfikacyjnym (ID) I0099 i opisał ją już w 2015 r. zespół Wolfganga Haaka, z tą różnicą, że jej wiek oszacowano wtedy na lata 1113 – 1021 p.n.e. [3]
ŁOTWA:
Ciekawa sytuacja przedstawia się w przypadku 15 próbek z Łotwy. Wszystkie pochodzą ze stanowiska Zvejnieki. Tutaj pojawiają się próbki o haplogrupie R1b (8 szt.; ok. 53%) oraz I2a (6 szt.; 40%). Oprócz nich jest tylko jedna egzotyczna próbka Q1a2 (7%). Co zaskakujące: najstarsza próbka to R1b1a1a datowana na lata 7465 – 7078 p.n.e. (krąg łowców-zbieraczy). 11 próbek ma taką samą datację na lata 6000 – 5100 p.n.e. (również łowcy zbieracze). Najmłodsze trzy próbki związane są już z ludnością neolityczną i wszystkie należą do hg R1b1a1a. Datowane są kolejno na lata: 5841 – 5636 p.n.e., 5302 – 4852 p.n.e. i 4251 – 3976 p.n.e. Sytuacja jest ciekawa z tego względu, gdyż obecnie na Łotwie obie haplogrupy (R1b i I2a) są nielicznie reprezentowane. Współcześni mieszkańcy Łotwy o hg R1b stanowią jedynie 12% męskiej ludności, zaś o hg I2a jedynie 1%! Dziś dominuje w tym kraju ludność o dwóch innych haplogrupach: R1a (40%) i N (38%). [4] Wynika z tego, że w pewnym okresie dziejów doszło do wymiany lub nawet kilku wymian ludności na terenie obecnych państw bałtyckich. Na pytanie kiedy to się stało postaram się odpowiedzieć w dalszej części tekstu, omawiając pracę niemieckiej genetyk Alissy Mittnik. [5]
BAŁKANY:
Podobnie jak na Łotwie, tak i również na Bałkanach ludność o haplogrupach R1b oraz I2a zamieszkiwała dość licznie w prehistorycznych czasach. Zespół Mathiesona opisał w swej pracy aż 58 próbek Y-DNA z terenu państw bałkańskich. Najstarsze 4 próbki pochodzą z miejscowości Padina w Serbii. Wśród nich są 3 próbki R1b (wszystkie R1b1a). Ich datowanie to kolejno lata: 9500 – 6200 p.n.e., 9221 – 8548 p.n.e., 8753 – 8351 p.n.e. Jedna próbka z czterech najstarszych to I2a1 datowana na 8753 – 8351 p.n.e. Na dziewiąte tysiąclecie datowana jest także próbka ze stanowiska Vlasac, również w Serbii. Jej wiek określa się na lata 8240 – 7940 r. p.n.e., a haplogrupę Y-DNA jako I.
Z okresu ósmego i siódmego tysiąclecia p.n.e. pochodzi 17 próbek bałkańskich. Ich dolna granica datowania wchodzi nieznacznie na początek szóstego tysiąclecia (7580 – 5841 r. p.n.e.). Próbki pochodzą z trzech państw: Serbii (12 szt.), Rumunii (3 szt.) i Bułgarii (2 szt.). Szczegóły ich haplogrup przedstawiają się następująco:
R1b1a – 5 szt.
I2a2a1b2 – 3 szt.
I2 – 2 szt.
C – 1 szt.
G2a2b2b1a – 1 szt.
I – 1 szt.
I2a2 – 1 szt.
I2a2a – 1 szt.
I2a2a1b – 1 szt.
R – 1 szt.;
Sprowadzając je do główniejszych haplogrup ich liczba i odsetek wyglądają tak:
I2a – 6 szt.; ok. 35% (3 x I2a2a1b2 + I2a2a1b + I2a2a + I2a2)
R1b – 5 szt.; ok. 29% (5 x R1b1a)
G2a – 1 szt.; ok. 6% (G2a2b2b1a)
etc.
Natomiast sprowadzając je do haplogrup oznaczonych tylko samymi pierwszymi literami wygląda to w ten sposób:
I – 9 szt.; ok. 53%
R – 6 szt.; ok. 35%
C – 1 szt.; ok. 6%
G – 1 szt.; ok. 6%
W niniejszym zestawieniu rzuca się w oczy brak hg R1a w ówczesnym czasie na Bałkanach, dziś dość licznie tam występującej.
Kolejne w chronologii, szóste tysiąclecie p.n.e. reprezentowane jest przez 13 próbek pochodzących z 5 państw – oprócz wcześniej wymienionych, dodatkowo z Chorwacji i Macedonii. Ich datowanie mieści się w przedziale 6000 – 5376 lat p.n.e. Zaskakujący jest brak nie tylko hg R1a, ale i R1b, wcześniej występującej w wyraźnym odsetku. Zmalał również odsetek hg I2a, zwiększyła się zaś liczba próbek hg G2a, wiązanej z ekspansją rolników z Bliskiego Wschodu i Anatolii. Oto szczegóły:
G2a – 6 szt.; ok. 46% (2 x G2a2b2a + G2a2a1 + G2a2b2b + G2a2b2b1 + G2a2a1a2a)
C + C1a2 – 3 szt.; ok. 23% (2x C + C1a2)
T1a – 2 szt.; ok. 15% (T1a1 + T1a1a)
E1b – 1 szt.; ok. 7,5% (E1b1b1a1b1)
I2a – 1 szt.; ok. 7,5% (I2a2)
Piąte tysiąclecie p.n.e. to 11 próbek z pracy Mathiesona et al. Pochodzą one z trzech państw: Bułgarii (8 szt.), Serbii (2 szt.) i Chorwacji (1 szt.). Ich datowanie rozpościera się od 4790 r. p.n.e. do 4264 r. p.n.e. W porównaniu do poprzedniego milenium, w tym okresie hg G2a nadal występuje najliczniej. Ponownie pojawia się R1b, która być może nieprzerwalnie występowała na Bałkanach, jednak stosownych próbek z okresu szóstego tysiąclecia nie znalazł zespół Mathiesona. Co dziwne – wciąż słabo występuje hg I, obecnie najliczniejsza na półwyspie Bałkańskim:
G2 + G2a – 5 szt.; ok. 46%
R + R1 + R1b – 3 szt.; 27%
CT – 2 szt.; 18 %
I – 1 szt.; 9%
Trzecie i czwarte milenium p.n.e. to 10 próbek z dwóch państw: Bułgarii (8 szt.) i Chorwacji (2 szt.). Tutaj nareszcie liczniej pojawia się hg I2a (5 szt.), dziś dominująca na Bałkanach, która zapewne występowała na Bałkanach nieprzerwanie od momentu pojawienia się tam (nawet 11.000 lat p.n.e. lub wcześniej). Nadal widoczna jest G2a oraz R1b:
I + I2a – 6 szt.; 60% (1 x I + 5 x I2a)
G2a – 2 szt.; 20%
H2 – 1 szt.; 10%
R1b – 1 szt.; 10%
Dwie najmłodsze bałkańskie próbki z niniejszej pracy naukowej pochodzą z drugiego tysiąclecia p.n.e. (1750 – 1500 r. p.n.e.). Nareszcie pojawia się R1a (dokładnie R1a1a1b2) wydobyta ze szkieletu w miasteczku Merichleri w Bułgarii (Merichleri – Kairyaka necropolis). Próbka ta oznaczana jest też jako R1a-Z93. Ta gałąź obecnie charakterystyczna jest dla mieszkańców Azji. Haplogrupa ta cechowała starożytnych Ariów, którzy przynieśli ją z Europy do Azji. [6][7]
Druga najmłodsza próbka to J2b2a z Chorwacji (Veliki Vanik).
TURCJA:
Obszar tureckiej Anatolii reprezentowany jest tylko przez jedną próbkę. Jest nią datowana na lata 2558 – 2295 p.n.e. (epoka brązu) próbka ze stanowiska Harmanoren-Gondurle Hoyuk. Jej haplogrupa nie budzi dużego zaskoczenia, gdyż jest nią J1a, współcześnie również obecna u męskiej populacji Turcji w 9%.
UKRAINA:
Na koniec zostawiłem przedstawienie kopalnych próbek Y-DNA z Ukrainy. Jest to ważna część niniejszego tekstu, gdyż związana jest z historią Słowian i związanej z nimi haplogrupą R1a. Praca naukowa „The Genomic History Of Southeastern Europe” zawiera aż 37 dawnych próbek z terytorium państwa ukraińskiego. W miejscowości Wasylewka (ang. Vasil’evka) odkryto jak na razie najstarszą próbkę R1a, datowaną na lata 8825 – 8561 p.n.e. Tym samym pod względem wieku zdetronizowana została do tej pory znana najstarsza próbka tej haplogrupy pochodząca ze stanowiska Jelenia Wyspa Południowa (ang. Yuzhnyy Oleni Ostrov) na Jeziorze Onega w rosyjskiej Karelii (Fu et al. 2016). [8]
Na Ukrainie udało się także wyodrębnić najstarszą próbkę subkladu M417 (R1a-M417 lub R1a1a1), który obejmuje prawie 100% współczesnych linii R1a. Próbka o numerze (ID) I6561 pochodzi ze stanowiska Alexandria i datowana jest na lata 5000 – 3500 p.n.e. Poniższa mapa pokazuje najstarsze gałęzie haplogrupy R1a:
– „Oldest R1a” to najstarsza do tej pory odkryta próbka R1a – Vasil’evka, Ukraina; ID I1819 (Mathieson et al. 2017),
– „Oldest R1a-M417” to najstarsza odkryta próbka R1a-M417 – Alexandria, Ukraina; ID I6561 (Mathieson et al. 2017),
– „Oldest R1a-Z93” to najstarsza odkryta próbka R1a-Z93 – Potapovka, obwód samarski, Rosja; ID 10432 / SVP42 (Mathieson 2015). [9]
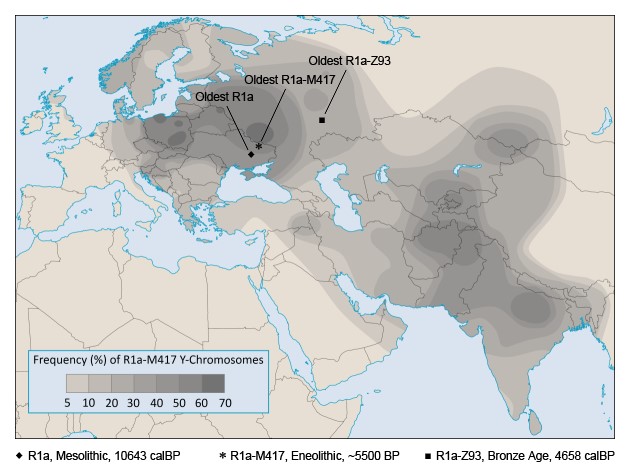
Rys. 1: Najstarsze odkryte próbki R1a, R1a-M417 i R1a-Z93; źródło: Eurogenes Blog.
Wynika z tego, że najstarsze miejsca zamieszkiwania ludności o hg R1a i jej gałęzi synowskich (okres 8500 – 5500 lat p.n.e.) to pas współczesnej Zachodniej Rosji i sąsiadujących z nią państw, rozciągający się na północy od Karelii po Wschodnią Ukrainę na południu. Być może kolejne badania pozwolą jeszcze rozszerzyć ten obszar.
Odkrycie starej próbki R1a-Z93 (Potapovka; ID 10432 / SVP42) na terenie Europy dowodzi, że gałąź Z93 narodziła się jeszcze w Europie, przed migracją ludności tego rodu do Azji. To właśnie ten subklad dominuje obecnie w Azji wśród głównie indoeuropejskiej ludności tego kontynentu. Ludność ta pochodzi od starożytnych Ariów, dalekich kuzynów Słowian, których migrację ze wschodnioeuropejskich stepów do Azji datuje się na okres 3000 – 1500 lat p.n.e. Sama R1a zrodziła się najprawdopodobniej w północnej Eurazji. Subklad R1a-M417 powstać musiał gdzieś na stepie nadczarnomorskim lub nadkaspijskim, na terenie dzisiejszej Ukrainy lub południowej Rosji. Miało to miejsce jakieś 7000 – 6000 lat temu. Nosiciele tego subkladu w przeciągu kilku tysięcy lat rozeszli się niemal we wszystkich kierunkach obejmując ogromny obszar od Skandynawii po Azję Południową. [10]
Należy wspomnieć, że oprócz próbki z Potapovki zespół Mathiesona w swej wcześniejszej pracy z 2015 r. odkrył inne próbki R1a-Z93 w europejskiej części Rosji. Wszystkie pochodzą z obwodu samarskiego. Są one nieco młodsze niż ta z Potapovki, bo datowane na lata 1900 – 1200 p.n.e. i związane są z kulturą grobów zrębowych (ang. Srubnaya culture). Oto one:
– Novosel’ki (ID 10232 / SVP12) – R1a1a1b2
– Barinovka I (ID 10423 / SVP31) – R1a1a1b2
– Uvarovka I (ID 10424 / SVP32) – R1a1a1b2
– Spiridonovka II (ID 10430 / SVP 39) – R1a1a1b2a2a. [9]
Wracając do tematu pozostałych próbek ukraińskich z pracy Mathiesona et al. 2017, obok wspomnianej próbki R1a z Wasylewki, kolejne dwie najstarsze również pochodzą z tego samego stanowiska. Pierwsza z nich zawiera haplogrupę I2a1 (ID I1763; 8280 – 7967 p.n.e.), zaś druga to próbka o hg R1b1a (ID I1734; 7446 – 7058 p.n.e.).
Cztery następne próbki datowane są na ten sam szeroki okres czasu 6500 – 4000 p.n.e. Wszystkie należą do haplogrupy I (3 x I, 1 x I2a2).
Aż 21 próbek posiada wspólne datowanie na lata 5500 – 4800 p.n.e. Dodając do nich jedną próbkę o zbliżonym datowaniu na lata 5473 – 5326 p.n.e. mamy wśród nich:
6 próbek I2a (I2a2, I2a2a, I2a2a1b, I2a2a1b1b, 2 x I2a2a1b1),
5 próbek R1b1a,
2 próbki I,
2 próbki R,
2 próbki R1,
1 próbka I2,
1 próbka IJ,
1 próbka R1a.
Zastanawia mała liczba R1a i dość spora R1b. Nie jest natomiast zaskoczeniem duża liczba próbek I2a oraz dodatkowo I oraz I2. Wśród współczesnych męskich mieszkańców Ukrainy najliczniej występuje hg R1a (44%), I2a (20,5%), R1b (8%) i E1b (6,5%). Zatem można ostrożnie mówić o pewnej kontynuacji zasiedlenia tego kraju od omawianego okresu po dzień obecny.
Kolejne 5 próbek z terenu Ukrainy pochodzą ze stanowiska kultury Cucuteni-Trypole, zlokalizowanego w jaskini Werteba (ang. Verteba Cave) w obwodzie tarnopolskim. Próbki łącznie datowane są na lata 4000 – 2936 p.n.e. Cztery z nich należą do hg G2a, zaś jedna do hg E.
Ostatnie dwie próbki związane są z kulturą amfor kulistych i obie należą do hg I2a (I2a2a1b i I2a2a1b2). Ich wiek oscyluje między 2899, a 2694 r. p.n.e.
MARK LIPSON Z ZESPOŁEM, 2017 r. – WĘGRY, NIEMCY, HISZPANIA
Z pracy, którą w marcu 2017 r. opublikował Mark Lipson z zespołem dowiedzieć się można o pochodzeniu wczesnych europejskich farmerów. [11] Z próbek zebranych na terenie Węgier, Niemiec i Hiszpanii wynika, iż w tych państwach w okresie między 5832, a 2212 r. p.n.e. dominowały haplogrupy pochodne od G oraz I. Szczegóły przedstawia poniższa tabelka: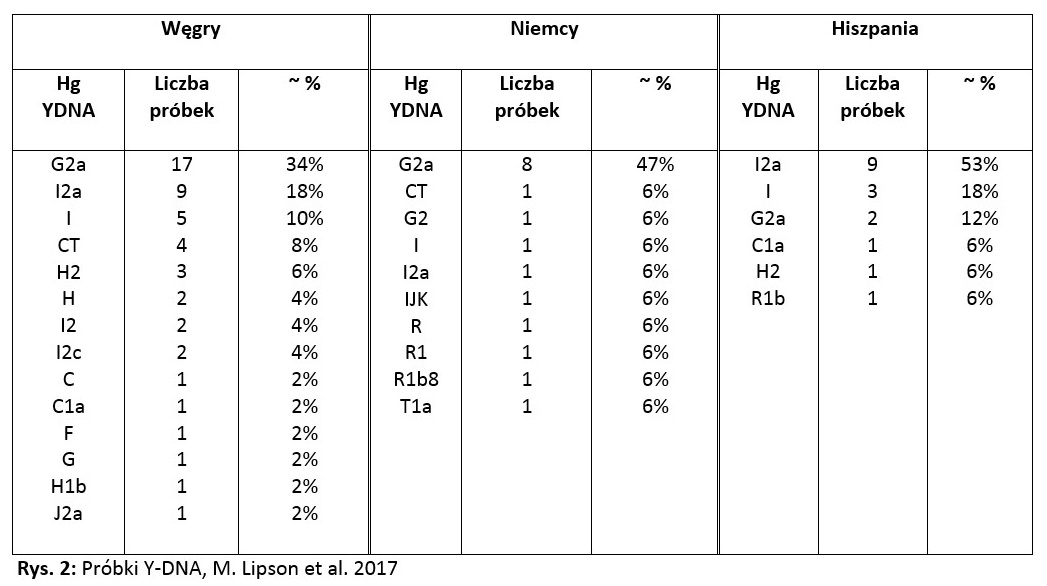
W przypadku Węgier, gdyby połączyć najliczniejsze haplogrupy G2a i G razem oraz I, I2a, I2 i I2c, to łączna liczba ich próbek oraz odsetek procentowy przedstawiłyby się następująco:
G: G2a + G = 18 próbek (36%),
I: I + I2 + I2a + I2c = 18 próbek (36%).
W przypadku Niemiec połączenie najliczniejszych haplogrup przedstawiałoby się tak:
G: G2 + G2a = 9 próbek (53%),
R: R + R1 + R1b = 3 próbki (18%),
I: I + I2a = 2 próbki (12%).
Analogiczny zabieg w przypadku Hiszpanii daje taki wynik:
I: I + I2a = 12 próbek (71%),
G: G2a = 2 próbki (12%).
Porównując te wyniki z haplogrupami współczesnych mieszkańców tych państw stwierdzić należy, że doszło w nich do wyraźnej wymiany ludności między omawianym w niniejszej pracy okresem, a czasami historycznymi. Między szóstym, a drugim tysiącleciem na Węgrzech dominowała ludność o hg G oraz I, znaczący odsetek stanowiła też hg H. Obecnie zaś dominują: R1a (29,5%), R1b (18,5%), I2a (16%), I1 (8,5%). Haplogrupa G wśród dzisiejszych Węgrów to zaledwie 3,5%, a hg H w ogóle nie jest podawana w statystykach, czyli praktycznie nie występuje. [4]
W przypadku Niemiec, sytuacja jest minimalnie lepsza, gdyż można mówić w jakimś stopniu o kontynuacji zamieszkania ziem niemieckich przez ludność o hg R (głównie R1b) oraz I. Natomiast ich proporcje uległy znacznym zmianom, bo obecnie sytuacja wygląda następująco: R1b (44,5%), R1a (16%), I1 (16%), E1b (5,5%), G (5%), I2b (4,5%), J2 (4,5%) oraz I2a (1,5%). Tutaj ponownie znacznie zmalał odsetek hg G, zwiększył się zaś odsetek hg R. W szczątkowej ilości zachowała się hg T (1%). [4]
Jeszcze bardziej drastycznie sytuacja przedstawia się w przypadku Hiszpanii. Z dominującej w odległych czasach haplogrupy I (71%) dziś pozostał ledwie 7%, a więc jej występowanie zmniejszyło się 10-krotnie. Dziś w tym kraju bezwzględnie dominuje hg R1b – obecnie 69%, a w omawianym okresie 6%. Haplogrupa G, niegdyś widoczna (12%), dziś stanowi tylko 3% wśród mieszkańców Hiszpanii. Natomiast haplogrupa H nie jest ujmowana w statystykach. [4]
W niniejszych badaniach istotna jest jeszcze jedna kwestia. Wiek próbek zawiera się w dość szerokim przedziale czasu między rokiem 5832 r., a 2212 r. p.n.e. Dlatego ważne jest określenie, które próbki były najstarsze w poszczególnych państwach. W przypadku Węgier są to dwie próbki H2 (5832 – 5536 p.n.e.) ze stanowiska Alsónyék-Bátaszék, Mérnöki telep, a w następnej kolejności próbki G2a (5800 – 5547 p.n.e.) ze stanowisk Lánycsók i ponownie Alsónyék-Bátaszék, Mérnöki telep. Tuż po G2a następuje próbka I2 datowana na lata 5302 – 5057 p.n.e. (stanowisko Hajdúnánás-Eszlári út).
Najstarsze próbki z terytorium Niemiec to 7 próbek G2a i jedna G2. Wszystkie one pochodzą ze stanowiska Halberstadt-Sonntagsfeld i są datowane na lata między 5500, a 4850 r. p.n.e. Najstarsza próbka R pochodzi z okresu 3654 – 3527 p.n.e. (Quedlinburg). Próbka R1b (R1b1) datowana zaś jest na lata 3958 – 3344 p.n.e. (Blätterhöhle). Zatem według tych wyników ludność o hg R / R1b pojawiła się na terenie Niemiec wyraźnie później niż ludność o hg G2a. Podobne wnioski wynikają z powyżej przedstawionych badań zespołu Mathiesona.
Najstarsza hiszpańska próbka to I2a (I2a1b1; 5310 – 5078 p.n.e.) ze stanowiska Els Trocs. Niewiele młodsza jest próbka R1b (R1b1a2; 5294 – 5066 p.n.e.) z tegoż samego stanowiska.
W odniesieniu do Niemiec i Hiszpanii, próbki przedstawione w pracy Lipsona (et al.) pokrywają się w niektórych przypadkach z próbkami, które w swej pracy przedstawił Wolfgang Haak z zespołem w 2015 r. [3]
Na podstawie badań zespołu Lipsona et al. wyłania się obraz, z którego wynika, iż europejskie populacje szóstego tysiąclecia p.n.e. pochodziły od imigrantów z Anatolii (hg G2a), którzy przybyli na europejskie ziemie zamieszkałe już przez łowców-zbieraczy (hg I1 + I2). Wbrew powyższym datowaniom uznać bowiem należy, że ludność łowców-zbieraczy (hg I1 + I2) była obecna w Europie jako jedna z pierwszych i tutaj jest jej kolebka. Wynika to z innych badań genetycznych. [12] [13]
Na podstawie poniższej mapy współczesnego rozmieszczenia hg G, można spekulować na temat domniemanej kolebki ludności o tej haplogrupie. Przyjmując za kolebkę miejsce największego zagęszczenia hg G, okazuje się, że mieściła się ona w rejonie południowego Kaukazu. Do dziś hg G występuje tam w największym odsetku. Dzięki mapie możemy określić domniemaną drogę tej ludności do Europy przez Anatolię, gdzie jej występowanie jest również dość wysokie, a następnie przez Bałkany. Istnieje hipoteza, że to właśnie ta ludność przyniosła do Europy rolnictwo. Biorąc pod uwagę fakt, iż systematyczna uprawa roli zaczęła się na Bliskim Wschodzie ok. 8000 lat p.n.e., koncepcja ta wydaje się być prawdopodobna. Kolejne badania pokażą czy jest ona w pełni słuszna.
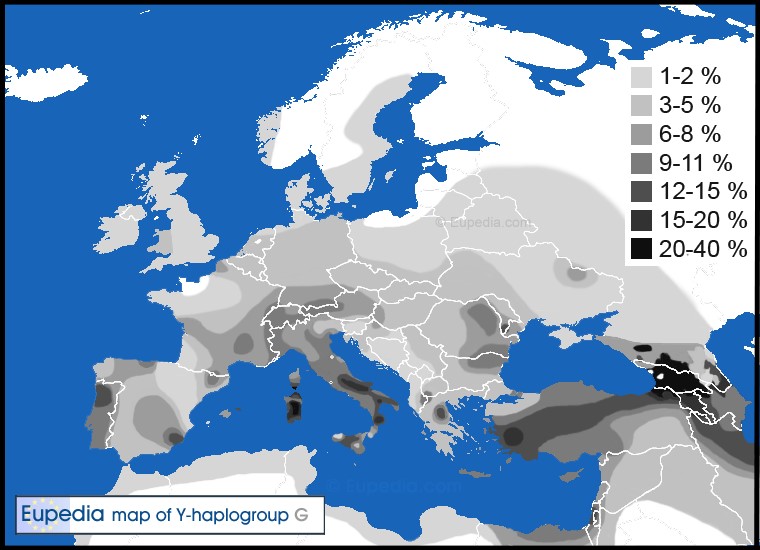
Rys. 3: Haplogrupa G; źródło: Eupedia.
ALISSA MITTNIK Z ZESPOŁEM, 2017 r. – ESTONIA, LITWA, ŁOTWA, SZWECJA
Zespół niemieckiej genetyk Alissy Mittnik postanowił zbadać obszar Europy mało jeszcze poznany pod względem genetycznym. Mowa o Europie Północnej, a zwłaszcza o wschodnim wybrzeżu Morza Bałtyckiego. [5] Terytorium te, jak i również Skandynawia zostało stopniowo zasiedlane 11.000 lat temu w miarę ustępowania lodowca. Analiza wieku i haplogrup próbek pozwala nam ponownie stwierdzić, że ludność łowców-zbieraczy (hg I1+I2) zamieszkiwała ziemie europejskie, a konkretnie północno-wschodniego wybrzeża Bałtyku jako jedna z pierwszych, o ile nie pierwsza. W pracy Alissy Mittnik najstarsze wyróżnione próbki Y-DNA posiadają hg I2a. Są to próbki należące do ludności tworzącej kulturę Narva. Kultura ta powstała na podłożu mezolitycznym wcześniejszej kultury kundajskiej. Należała do kultur subneolitycznych strefy leśnej. Na podstawie artefaktów stwierdzono, że jej ludność zajmowała się głównie polowaniami, rybołówstwem i zbieractwem. Dwie wyróżnione próbki tej kultury pochodzą z Litwy: jedna pochodząca ze stanowiska przy jeziorze Kretuonas i druga ze stanowiska na wyspie Spiginas na jeziorze Biržulis. Pierwsza datowana jest na lata 5500 – 2900 p.n.e., zaś druga na 4440 – 4240 p.n.e.
Dopiero około połowy trzeciego tysiąclecia na ziemiach obecnych państw bałtyckich masowo pojawia się ludność indoeuropejska o haplogrupie R1a. Najstarsza próbka tej haplogrupy wyodrębniona przez zespół A. Mittnik datowana jest na lata 2620 – 2470 (stanowisko Gyvakarai, Litwa).
Szczegóły przedstawia poniższa tabela (Rys. 4):
Wniosek wynikający z wyników zawartych w tabeli to znaczny wysyp próbek haplogrupy R1a począwszy od trzeciego tysiąclecia p.n.e. Podobne wnioski na temat tej haplogrupy można wysnuć z dwóch poprzednich prac, jakie ukazały się dwa lata wcześniej, a więc w 2015 r. Mam tu na myśli wspomnianą już powyżej pracę zespołu Wolfganga Haaka oraz pracę zespołu Mortena E. Allentofta. [3] [14]
W pracy A. Mittnik wyróżnia się próbka R1a ze Szwecji (stanowisko Ölsund). Wspomniany M. E. Allentoft także wyróżnił jedną próbkę R1a ze Szwecji datowaną na podobny czas, bo na 2550 r. p.n.e. (stanowisko Viby). Na ten okres w południowej części Skandynawii przypada kultura ceramiki sznurowej kojarzona właśnie głównie z ludnością rodu R1a, a zarazem z ludnością prasłowiańską. To właśnie w tym czasie według wszelkich danych nastąpiła ekspansja tejże ludności w różnych kierunkach Europy i Azji, o czym miałem już okazję pisać w innym artykule. [6]
Allentoft wyodrębnił 6 próbek Y-DNA z okresu kultury ceramiki sznurowej, z czego 5 to próbki R1a. Przedstawia to poniższa tabela (Rys. 5):
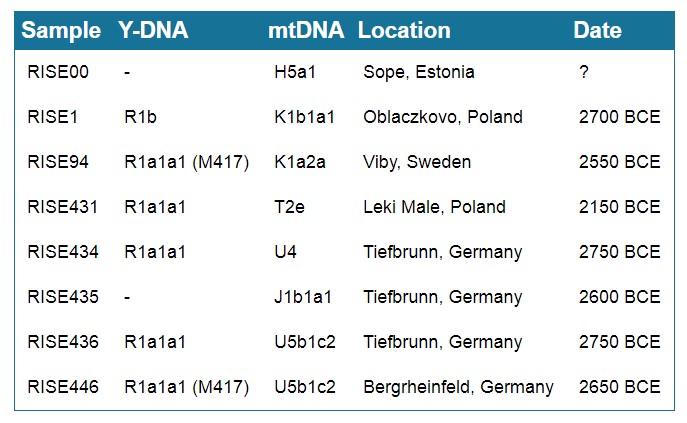
Rys. 5: Próbki Y-DNA i mtDNA z kręgu kultury ceramiki sznurowej, M. E. Allentoft et al. 2015,
źródło: Eupedia.
- Haak natomiast, wyodrębnił tylko jedną próbkę Y-DNA z tejże kultury datowaną na lata 2473 – 2348 p.n.e. (stanowisko Esperstedt, Niemcy). Była to także próbka o haplogrupie R1a, a dokładnie hg R1a1a1. Oprócz niej, Haak wyodrębnił inne próbki datowane na okres trzeciego tysiąclecia p.n.e. Oto one:
– R1b1a2a1a2 – Quedlinburg, Niemcy (2296 – 2206 p.n.e.), k. pucharów dzwonowatych,
– I2 – Eulau, Niemcy (2131 – 1982 p.n.e.), k. unietycka,
– I2a2 – Esperstedt, Niemcy (2131 – 1979 p.n.e.), k. unietycka,
– I2c2 – Esperstedt, Niemcy (2118 – 1961 p.n.e.), k. unietycka.
W ramach krótkiego komentarza zwracam uwagę, że występowanie haplogrupy R1b u ludności kultury pucharów dzwonowatych jest powszechne, o czym będzie mowa w kolejnym artykule (część 2). Również znalezienie próbki o hg R1b w miejscowości Obłaczkowo (woj. wielkopolskie, pow. wrzesiński) na stanowisku kultury ceramiki sznurowej, zdominowanej przez ród R1a, dowodzi, iż hg R1b obecna była na terenie Polski na długo przed średniowiecznym niemieckim i ogólnie zachodnioeuropejskim osadnictwem. Ten temat także będzie poruszony we wspomnianym, kolejnym artykule.
Natomiast rody I2 oraz I1, o czym już wspominałem, są obecne w wielu dawnych kulturach w różnych miejscach, niemal całej Europy. Powtórzę: haplogrupy I1 oraz I2 zrodziły się najprawdopodobniej w Europie i najprawdopodobniej zasiedliły rozległe jej terytoria jako pierwsze. A w każdym razie na pewno wcześniej niż rody R1a i R1b.
Haplogrupa I również współcześnie jest mocno rozpowszechniona w Europie. W niemałym odsetku spotykana jest w różnych państwach. Dominuje na Bałkanach (Bośnia i Hercegowina: 50,5% I2 + 4,5% I1) oraz w Skandynawii (Szwecja: 37% I1 + 5% I2). Wśród współczesnych Polaków haplogrupa I jest na drugim miejscu, zaraz po haplogrupie R (R1a + R1b) i wynosi łącznie 16%, z czego 8,5% to I1, zaś 7,5% to I2. [4]
Porównując próbki z pracy zespołu I. Mathiesona z próbkami ukazanymi w pracy zespołu Alissy Mittnik daje się zauważyć wymiana ludności na terenie wschodniego wybrzeża Bałtyku, jaka nastąpiła najprawdopodobniej w czwartym tysiącleciu p.n.e. Przypomnijmy: w szóstym i piątym tysiącleciu p.n.e. na terenie Łotwy zidentyfikowano kopalne Y-DNA należące głównie do rodu R1b oraz I2a (łącznie 93% wszystkich odkrytych próbek; Mathieson et al. 2017). Natomiast próbki zidentyfikowane przez A. Mittnik et al. pochodzą głównie z okresu od trzeciego tysiąclecia do III w. p.n.e. i wszystkie należą do rodu R1a. Dwie starsze próbki z okresu 5500 – 2900 to próbki I2a.
Kolejnym zaskakującym odkryciem jest fakt, że ani w pracy zespołu Mathiesona, ani w pracy zespołu Mittnik nie wyróżniono żadnej próbki o hg N. Jest to o tyle dziwne, gdyż obecnie w państwach bałtyckich i w Finlandii męska część populacji o tej haplogrupie jest najbardziej liczna (Finlandia 61,5%, Litwa 42%, Estonia 34%) lub nieznacznie ustępuje R1a (Łotwa: 40% R1a i 38% N). Świadczy to najprawdopodobniej o tym, że po okresie kultury ceramiki sznurowej na teren wschodniego wybrzeża Bałtyku przybyła kolejna fala migracyjna – tym razem ludności o hg N. Przybyła ona ze wschodu, z okolic Uralu. W państwach takich jak Finlandia i Estonia język tej ludności wyparł wcześniejsze języki, w tym indoeuropejskie, zaś na Litwie, Łotwie i w Prusach ludność ta najprawdopodobniej doprowadziła do wykształcenia się nowego etnosu – Bałtów i tym samym nowej grupy językowej – bałtyckiej, która oderwała się od swego prasłowiańskiego rdzenia i która zarazem zachowała indoeuropejski charakter swojej mowy. Potwierdziłoby to, co miałem okazję już przedstawić w innych artykułach. [6] [17]
LEHTI SAAG Z ZESPOŁEM, 2017 r. – ESTONIA
Wnioski płynące z analizy próbek zawartych w pracy Alissy Mittnik et al. potwierdza praca estońskich naukowców pod kierownictwem Lehti Saag. [15] Dotyczy to wymiany ludności, która dokonała się między szóstym, a trzecim tysiącleciem p.n.e. – jak już wspomniałem – najprawdopodobniej około czwartego tysiąclecia p.n.e.
Praca, którą opublikowała Lehti Saag pokazuje, że w tym okresie nastąpiła dość wyraźna zmiana męskiej populacji na terenie współczesnych państw wschodniego wybrzeża Bałtyku. Tak jak wcześniej dominowała tu ludność o hg R1b (Łotwa – Mathieson et al. 2017), tak w pracy Saag et al. 2017 dominuje ludność o hg R1a (Estonia). Co jeszcze bardziej zaskakujące – w badanym przez zespół estońskiej genetyk okresie brak jest próbek o hg N, a więc tej, która dziś jest najliczniejsza w Estonii (34%). Konkluzje z tego są następujące i pokrywają się z konkluzjami zawartymi powyżej przy omawianiu pracy A. Mittnik et al.:
- Okres kultury ceramiki sznurowej to silna ekspansja ludności z rodu R1a, która wyparła z terenu wschodniego Bałtyku wcześniejszą ludność, głównie męską (głównie o hg R1b i I2a).
- Potwierdzają się wcześniejsze poglądy, że ludność o hg N (ugrofińska) przybyła do Estonii i Finlandii oraz na Łotwę i Litwę stosunkowo późno. W okresie kultury ceramiki sznurowej nie była jeszcze tam obecna.
Odnośnie punktu 2. warto wspomnieć co na temat przybycia ludności fińskiej i lapońskiej napisał fiński naukowiec Mikko K. Heikkilä w swej książce pt. „Bidrag till Fennoskandiens språkliga förhistoria i tid och rum” o językowej prehistorii „Fennoskandii”. [16] Autor uważa, że językowi przodkowie Finów i Lapończyków (Samów) przybyli w regiony północnego i wschodniego wybrzeża Bałtyku na początku epoki brązu z dorzecza Wołgi i Kamy. Wcześniej na terenie Finlandii i Estonii mieszkała ludność indoeuropejska i paleoeuropejska. Mimo, iż fiński naukowiec nie wspomina o Prasłowianach na tym terenie, to jednak w odniesieniu do Estonii można wysnuć wniosek, iż na Prasłowian obecnych tam od co najmniej epoki kultury ceramiki sznurowej (hg R1a) nałożyła się ludność ugrofińska o haplogrupie N (N1c). Na terenie Estonii ludność ta narzuciła swój język tamtejszym Prasłowianom o hg R1a, natomiast na terenie Łotwy i Litwy doszło do nałożenia się języka pra-ugrofińskiego na substrat języka prasłowiańskiego tworząc grupę języków bałtyckich. Języki te, w przeciwieństwie do języka estońskiego pozostały w rodzinie języków indoeuropejskich. W kwestii szczegółów niezbędne są jednak wnikliwe badania lingwistyczne.
W przeciwieństwie do książki Mikko K. Heikkilä, w pracy A. Mittnik przyjmuje się, że przybycie ludności ugrofińskiej o hg N na teren współczesnej Finlandii, Estonii, Łotwy czy Litwy miało miejsce pod koniec epoki brązu, a nie na jej początku. Osobiście skłaniam się bardziej ku tezie niemieckiej genetyk.

ZAKOŃCZENIE
Na podstawie krótkiego omówienie powyższych prac naukowych, a konkretnie próbek Y-DNA zawartych w tych pracach, stwierdzić należy, że badania genetyków wniosły, stale wnoszą i przez długi czas jeszcze wnosić będą ogrom informacji w kwestii pochodzenia Europejczyków. Wiedza ta jest wraz z każdą kolejną pracą naukową na bieżąco aktualizowana i uzupełniana. Jednocześnie upadają mity na temat pochodzenia licznych ludów europejskich, które oparte były o naiwne i tendencyjne interpretacje, nierzadko zaangażowanych politycznie naukowców, którzy pozbawieni byli nowoczesnego warsztatu naukowego, jaki dostępny jest dziś.
Kończąc ten artykuł zapraszam na kolejną część niniejszego zagadnienia, która obecnie (listopad 2017 r.) jest w fazie tworzenia. Kolejny artykuł z tej serii bardziej będzie związany z ziemiami polskimi oraz z pochodzeniem Słowian i Polaków.
Adrian Leszczyński
aleszczynski@interia.pl
PRZYPISY:
[1] – Iain Mathieson et al.; The Genomic History Of Southeastern Europe; bioRxiv 09.05.2017; https://www.biorxiv.org/content/early/2017/09/19/135616.full.pdf+html
[2] – Witold Hensel; Polska starożytna; Wrocław 1988, s. 130.
[3] – Wolfgang Haak et al.; Massive migration from the steppe is a source for Indo-European languages in Europe; bioRxiv 10.02.2015; https://www.biorxiv.org/content/biorxiv/early/2015/02/10/013433.full.pdf
[4] – źródło: Eupedia; https://www.eupedia.com/europe/european_y-dna_haplogroups.shtml
[5] – Alissa Mittnik et al.; The Genetic History of Northern Europe; bioRxiv 03.03.2017; https://www.biorxiv.org/content/early/2017/03/03/113241
[6] – Adrian Leszczyński; Krótka historia rodu genetycznego R1a; Białczyński 17.01.2017; https://bialczynski.pl/2017/01/17/adrian-leszczynski-krotka-historia-rodu-genetycznego-r1a-2/
– Skribh 17.01.2017; https://skribh.wordpress.com/2017/01/17/400-skrbh-11-adrian-leszczynski-krotka-historia-rodu-genetycznego-r1a/
[7] – Adrian Leszczyński; Języki słowiańskie a haplogrupa R1a1; Taraka 16.03.2016;
http://www.taraka.pl/jezyki_slowianskie_a_haplogrupa_R1a1
[8] – Qiaomei Fu et al.; The genetic history of Ice Age Europe; Nature 02.05.2016;
http://www.nature.com/nature/journal/v534/n7606/full/nature17993.html
[9] – Iain Mathieson et al.; Eight thousand years of natural selection in Europe; bioRxiv 14.03.2015;
https://www.biorxiv.org/content/early/2015/10/10/016477.full.pdf+html
[10] – Davidski; The beast among Y-haplogroups; Eurogenes Blog 26.09.2017;
http://eurogenes.blogspot.com.au/2017/09/the-beast-among-y-haplogroups.html
[11] – Mark Lipson et al.; Parallel ancient genomic transects reveal complex population history of early European farmers; bioRxiv 06.03.2017; http://biorxiv.org/content/early/2017/03/06/114488
[12] – Maciamo Hay; Haplogroup I2 Y-DNA; Eupedia 2017;
https://www.eupedia.com/europe/Haplogroup_I2_Y-DNA.shtml
[13] – Eppie R. Jones et al.; Upper Palaeolithic genomes reveal deep roots of modern Eurasians; Nature 16.11.2015;
https://www.nature.com/articles/ncomms9912
[14] – Morten E. Allentoft et al.; Population genomics of Bronze Age Eurasia; Nature 10.06.2015; http://www.nature.com/nature/journal/v522/n7555/full/nature14507.html
[15] – Lehti Saag et al.; Extensive farming in Estonia started through a sex-biased migration from the Steppe; bioRxiv 02.03.2017; https://www.biorxiv.org/content/early/2017/03/02/112714.full.pdf+html
[16] – Mikko K. Heikkilä; Bidrag till Fennoskandiens språkliga förhistoria i tid och rum; Helsinki 2014; https://helda.helsinki.fi/bitstream/handle/10138/135714/bidragti.pdf?sequence=1 (PDF w j. szwedzkim)
– artykuł w j. angielskim nt książki: Dienekes’ Antropology Blog 17.08.2014; http://dienekes.blogspot.com/2014/08/indo-europeans-preceded-finno-ugrians.html
[17] – Adrian Leszczyński; Języki indoeuropejskie a genetyka; Taraka 04.05.2014; http://www.taraka.pl/jezyki_indoeuropejskie_a_genetyka