Dynamic changes in genomic and social structures in third millennium BCE central Europe (Dynamiczne zmiany w strukturach genomowych i społecznych w trzecim tysiącleciu p.n.e. w Europie Środkowej)
Europe’s prehistory oversaw dynamic and complex interactions of diverse societies, hitherto unexplored at detailed regional scales. Studying 271 human genomes dated ~4900 to 1600 BCE from the European heartland, Bohemia, we reveal unprecedented genetic changes and social processes. Major migrations preceded the arrival of “steppe” ancestry, and at ~2800 BCE, three genetically and culturally differentiated groups coexisted. Corded Ware appeared by 2900 BCE, were initially genetically diverse, did not derive all steppe ancestry from known Yamnaya, and assimilated females of diverse backgrounds. Both Corded Ware and Bell Beaker groups underwent dynamic changes, involving sharp reductions and complete replacements of Y-chromosomal diversity at ~2600 and ~2400 BCE, respectively, the latter accompanied by increased Neolithic-like ancestry. The Bronze Age saw new social organization emerge amid a ≥40% population turnover.
Abstrakt -Skrót
Prehistoria Europy przechodziła dynamiczne i złożone interakcje różnych społeczeństw, dotychczas niezbadane szczegółowo w skali regionalnej.Badając 271 ludzkich genomów datowanych na ok. 4900-1600 p.n.e. z serca Europy, Czech, odkrywamy bezprecedensowe zmiany genetyczne i procesy społeczne.Duże migracje poprzedziły przybycie przodków „stepowych”, a około 2800 p.n.e. współistniały trzy grupy zróżnicowane genetycznie i kulturowo.Wyroby Kultury Ceramiki Sznurowej pojawiły się w 2900 r. p.n.e., początkowo grupy Sznurowców były zróżnicowane genetycznie, nie wywodziły się w całości z przodków stepowych znanych jako Kultura Yamnaya i zasymilowały kobiety o różnym pochodzeniu.Zarówno grupy Corded Ware (Kultury Ceramiki Sznurowej), jak i Bell Beaker (Kultury Pucharów Dzwonkowatych) przeszły dynamiczne zmiany, obejmujące gwałtowne redukcje i całkowite zastąpienie różnorodności chromosomu Y odpowiednio w ~2600 i ~2400 p.n.e., przy czym tym ostatnim towarzyszyły zwiększone migracje przypominające początek neolitu.W epoce brązu pojawiła się nowa organizacja społeczna przy co najmniej 40% rotacji populacji.
INTRODUCTION – Wstęp
Archaeogenetics has revealed two major population turnovers in Europe within the past 10,000 years (1–5). The first, beginning in the seventh millennium BCE, was associated with expanding Neolithic farming communities from Anatolia (6, 7). European Early Neolithic farmers were initially genetically distinct from preceding hunter-gatherers (HG) and almost indistinguishable from Anatolian farmers (8–10), however incorporated HG ancestry into their gene pools over ensuing millennia (3, 11–13).
Archeogenetyka ujawniła dwie główne rotacje populacji w Europie w ciągu ostatnich 10 000 lat (1–5).Pierwsza, zapoczątkowana w siódmym tysiącleciu p.n.e., związana była z rozrastaniem się neolitycznych społeczności rolniczych z Anatolii (6, 7). Kultury Europejskich Rolników z wczesnego neolitu były początkowo genetycznie różne od poprzedzających je kultur myśliwych i zbieraczy (HG – hunters and gatherers = Myśliwi i Zbieracze) i prawie nie do odróżnienia od rolników z Anatolii (8-10), jednak przez kolejne tysiąclecia włączały one osoby o pochodzeniu HG do swoich puli genów (3, 11-13).
The second major turnover occurred in the early third millennium BCE with individuals of the Corded Ware (CW) culture (3, 4, 8). Of note, in what follows, we use the co-occurrence of human skeletal remains and markers of archaeological cultures (e.g., grave goods and body orientation) to denote an association between individuals and an archaeological culture (e.g., “CW individuals”), although this may not reflect a unified social entity. The CW represents a major cultural shift in central, northern, and northeastern Europe, bringing changes in economy, ideology, and mortuary practices (14–22). CW individuals were shown to be genetically distinct from culturally pre-CW people, having ~75% of their ancestry similar to Yamnaya individuals from the Pontic-Caspian steppe (3, 4, 23–27). This Yamnaya-like “steppe” ancestry then spread rapidly throughout Europe, reaching Britain, Ireland, the Iberian Peninsula, the Balearic Islands, Sardinia, and Sicily before the end of the third millennium BCE (5, 28–32).
Druga znacząca zmiana nastąpiła na początku trzeciego tysiąclecia p.n.e. z pojawieniem się społeczności kultury ceramiki sznurowej (CW) (3, 4, 8). Warto zauważyć, że poniżej używamy współwystępowania ludzkich szczątków szkieletowych i znaczników kultur archeologicznych (np. dobra grobowe i orientacja ciała) w celu oznaczenia związku między jednostkami a kulturą archeologiczną (np. „osoby CW”), chociaż może to nie odzwierciedlać jednolitej jednostki społecznej. CW reprezentuje poważną zmianę kulturową w środkowej, północnej i północno-wschodniej Europie, przynosząc zmiany w ekonomii, ideologii i praktykach pogrzebowych (14-22). Wykazano, że osoby CW różnią się genetycznie odspołeczności kulturowych sprzed CW, mając ~75% przodków podobnych dospołeczności Yamnaya ze stepu pontyjsko-kaspijskiego (3, 4, 23-27). Ten podobny do Yamnaya „stepowy” przodek szybko rozprzestrzenił się w całej Europie, docierając do Wielkiej Brytanii, Irlandii, Półwyspu Iberyjskiego, Balearów, Sardynii i Sycylii przed końcem trzeciego tysiąclecia pne (5,28–32).
Despite the importance of the third millennium BCE, our genetic understanding is mainly built upon studies with pan-European sampling strategies, with little emphasis on regional, high-resolution temporal transects (3–5, 8). Consequently, many temporal and geographic sampling gaps remain, resulting in limited knowledge about the processes at the level of the societies and communities and how cultural groups interacted, influenced, and gave rise to one another. In addition, the use of small sample sizes to represent supra-regional archaeological phenomena, as well as the resulting oversimplified culture-historical interpretations, has drawn criticisms from archaeologists (21, 33–40).
Pomimo wiedzy o znaczeniu trzeciego tysiąclecia pne, nasze zrozumienie genetyczne opiera się głównie na badaniach z paneuropejskimi strategiami pobierania próbek, z niewielkim naciskiem na regionalne transekty czasowe o wysokiej rozdzielczości (3-5, 8).W konsekwencji pozostaje wiele czasowych i geograficznych luk w próbkach, co skutkuje ograniczoną wiedzą na temat procesów zachodzących na poziomie społeczeństw i społeczności oraz tego, w jaki sposób grupy kulturowe wchodziły w interakcje, wpływały na siebie nawzajem i tworzyły się nawzajem.Ponadto użycie niewielkich liczebnie próbek do prezentowania ponadregionalnych zjawisk archeologicznych, a także wynikające z nich nadmiernie uproszczone interpretacje kulturowo-historyczne, wywołały krytykę ze strony archeologów (21, 33–40).
Unresolved questions concern the genetic and geographic origins of CW and Bell Beaker (BB) individuals, their relationship to one another and to Yamnaya individuals, as well as the origin of Early Bronze Age (EBA) Únětice individuals. Although it has been proposed that CW formed from a male-biased westward migration of genetically Yamnaya-like people (23, 41–44), no overlap in Y-chromosomal lineages (with the exception of a few nondiagnostic I2) has been found between the predominantly R1a-carrying CW and mainly R1b-Z2103–carrying Yamnaya males. Steppe ancestry is also present in BB individuals (5); however, they predominantly carry R1b-P312, a Y-lineage not yet found among CW or Yamnaya males. Therefore, despite their sharing of steppe ancestry (3, 4) and substantial chronological overlap (45), it is currently not possible to directly link Yamnaya, CW, and BB groups as paternal genealogical sources for one another, particularly noteworthy in light of steppe ancestry’s suggested male-driven spread (23, 41–43) and the proposed patrilocal/patriarchal social kinship systems of these three societies (46–48).
Nierozwiązane pytania dotyczą genetycznego i geograficznego pochodzenia osobników CW i Bell Beaker (BB), ich relacji między sobą oraz z osobnikami Yamnaya, a także pochodzenia osobników z wczesnej epoki brązu (EBA) Únětice. Chociaż zaproponowano, że CW powstało z migracji ukierunkowanej na zachód przez ludzi genetycznie podobnych do Yamnaya (23, 41-44), nie stwierdzono nakładania się linii chromosomów Y (z wyjątkiem kilku niediagnostycznych I2) między głównie CW przenoszącym R1a i głównie mężczyznami Yamnaya przenoszącymi R1b-Z2103. Pochodzenie stepowe jest również obecne u osobników BB (5); jednak są oni głównie nosicielami R1b-P312, linii Y, której jeszcze nie znaleziono wśród mężczyzn CW lub Yamnaya. Dlatego pomimo ich wspólnego pochodzenia stepowego (3, 4) i znacznego nakładania się chronologicznego (45), obecnie nie jest możliwe bezpośrednie powiązanie grup Yamnaya, CW i BB jako ojcowskich źródeł genealogicznych dla siebie, co jest szczególnie godne uwagi w świetle historii kultur stepowych, gdzie sugerowano jako przodków rozprzestrzeniające się grupy mężczyzn (23, 41–43) oraz proponowane patrilokalne/patriarchalne wspólne systemy pokrewieństwa społecznego tych trzech społeczeństw (46–48).
Crucial to understanding the cultural, social, and genetic transitions in third millennium BCE Europe are densely settled regions that attest to the (co)existence of societies attributed to pre-CW [Baden and Globular Amphora (GAC)], CW, BB, and EBA Únětice. Currently, no such region has been systematically studied from the archaeogenetic perspective. Situated in the heart of Europe and tightly nestled around the important Elbe river, the fertile lowlands of Bohemia, the western part of today’s Czech Republic, witnessed many major supra-regional archaeological phenomena (table S1, Fig. 1, fig. S1, and the Supplementary Materials). Dense agrarian settlement of Bohemia began after ~5400 BCE (49, 50) with the arrival of early Neolithic farmers (Linearbandkeramik-LBK, later Stichbandkeramik-STK, and Lengyel). They were succeeded by manifold societies of the Eneolithic (~4400 to 2200 BCE), associated with more than a dozen archaeological cultural groups including Jordanów, Michelsberg, Funnelbeaker, Baden, Řivnáč, GAC, Early and Late CW, and BB (table S1) (50). The Eneolithic witnessed important innovations (metallurgy, the wheel, wagon and plough, fortified hillforts, and burial mounds) (51–53) and was succeeded by the globalized EBA Únětice culture, geographically centered around Bohemia.
Kluczowe dla zrozumienia przemian kulturowych, społecznych i genetycznych w trzecim tysiącleciu p.n.e. jest to, że Europa to gęsto zaludnione regiony, które świadczą o (współ)istnieniu społeczeństw przypisywanych jako pre-Sznurowe jak Kultura Badeńska i Kultura Amfor Kulistych (GAC) oraz CW, BB i EBA Únětice. Dotychczas żaden taki region nie był systematycznie badany z perspektywy archeogenetycznej. Położone w sercu Europy i ciasno zagnieżdżone wokół ważnej rzeki Łaby, żyzne niziny Czech, zachodnia część dzisiejszych Czech, były świadkiem wielu ważnych ponadregionalnych zjawisk archeologicznych (tabela S1, ryc. 1, ryc. S1 i Materiały Uzupełniające). Gęste osadnictwo rolnicze w Czechach rozpoczęło się po ~5400 pne (49, 50) wraz z przybyciem wczesnych rolników neolitycznych (kultury Linearbandkeramik-LBK, później Stichbandkeramik-STK i Lengyel). Zostały zastąpione przez różnorodne ugrupowania eneolitu (~4400 do 2200 pne), związane z kilkunastu archeologicznymi grupami kulturowymi, w tym Jordanów, Michelsberg, Funnelbeaker, Baden, Řivnáč, GAC, wczesne i późne CW oraz BB (tabela S1) (50). Eneolit był świadkiem ważnych innowacji (metalurgia, koło, wóz i pług, ufortyfikowane grodziska i kurhany) (51–53), a jego następcą została zglobalizowana kultura EBA Únětice, geograficznie skupiona wokół Czech.
Fig. 1. Temporal and geographic distribution of studied Neolithic, Eneolithic, and EBA individuals from Bohemia.
(A) Map of Bohemia showing the locations of sampled sites (red, new; blue, previously published; table S2 and figs. S1 to S5). (B) Mean age of newly reported (n = 206) and published (n = 65) individuals from Bohemia. (C) Local chronology of archaeological cultures and time periods. Black triangles indicate external influences visible in the material culture. Red lines indicate qualitative degree of change in material culture.
Ryc. 1. Rozmieszczenie czasowe i geograficzne badanych osobników neolitu, eneolitu i EBA z Czech.(A) Mapa Czech pokazująca lokalizacje pobranych miejsc (czerwona, nowa; niebieska, poprzednio opublikowana; tabela S2 i rys. S1 do S5).(B) Średni wiek nowo zgłoszonych (n = 206) i opublikowanych (n = 65) osób z Czech.(C) Lokalna chronologia kultur archeologicznych i okresów.Czarne trójkąty wskazują na zewnętrzne wpływy widoczne w kulturze materialnej.Czerwone linie wskazują jakościowy stopień zmian w kulturze materialnej.
In addition to material and technological developments, ideological changes, as manifested through mortuary behavior, are also evident (54). Although relatively common during the Funnelbeaker period (~3800 to 3400 BCE, n = ~100 known graves in Bohemia) (55), regular graves almost disappear from the succeeding Baden, Řivnáč, and GAC periods (Middle Eneolithic, ~3500 to 2800 BCE, n = ~20 in Bohemia) (56). Single graves, but now with strict gender differentiation in body position and grave goods, reappeared in abundance with CW from ~2900 BCE (n = ~1500 in Bohemia) (50, 57) and continued with BB (n = ~600 in Bohemia) from ~2500 BCE (58), who developed and maintained important differences from the preceding CW. The EBA Únětice culture (59, 60) continued with single graves (n = ~4000 to 5000 in Bohemia), but now again without gender differentiation in body position.
Oprócz rozwoju materialnego i technologicznego widoczne są również zmiany ideologiczne, przejawiające się w zachowaniach pogrzebowych (54).Chociaż stosunkowo powszechne w okresie pucharów lejkowatych (~3800 do 3400 pne, n = ~100 znanych grobów w Czechach) (55), regularne groby prawie znikają z kolejnych okresów Baden, Řivnáč i GAC (średni eneolit, ~3500 do 2800 pne, n = ~20 w Czechach) (56).Pojedyncze groby, ale teraz ze ścisłym zróżnicowaniem płci w pozycji ciała i dobrach grobowych, pojawiły się obficie z CW od ~2900 p.n.e. (n = ~1500 w Czechach) (50, 57) i były kontynuowane z BB (n = ~600 w Czechach)od ~2500 p.n.e. (58), który rozwinął i utrzymał istotne różnice w stosunku do poprzedniego CW.Kultura EBA Únětice (59, 60) kontynuowała je w pojedynczych grobach (n = ~ 4000 do 5000 w Czechach), ale teraz znowu bez różnicowania płci w pozycji ciała.
To better understand these transitions, we analyzed a high-resolution archaeogenetic time transect of 271 (206 newly reported and 65 previously published) individuals (Fig. 1, fig. S1, tables S2 to S4, and the Supplementary Materials) from the northern part of Bohemia. Through dense genetic sampling from geographically and temporally overlapping archaeological cultures, we aim to (i) address whether cultural changes in the Eneolithic and EBA of central Europe were driven by an influx of nonlocals, (ii) characterize the central European genetic diversity immediately prior the appearance of CW, (iii) date when individuals with Yamnaya-like steppe ancestry first appeared in central Europe and understand their genetic origin and social structure, (iv) characterize the nature and extent of biological exchange between the “locals” and “migrants” after the appearance of CW, and (v) identify social transformations linked to genetic and archaeological changes.
Aby lepiej zrozumieć te przejścia, przeanalizowaliśmy wysokiej rozdzielczości archeogenetyczny transekt czasu obejmujący 271 (206 nowo zgłoszonych i 65 wcześniej opublikowanych) osobników (ryc. 1, ryc. S1, tabele od S2 do S4 oraz materiały dodatkowe) z północnej częściCzech.Poprzez gęste pobieranie próbek genetycznych z geograficznie i czasowo nakładających się kultur archeologicznych, staramy się:
(i) ustalić, czy zmiany kulturowe w eneolicie i EBA w Europie Środkowej były spowodowane napływem grup nielokalnych,
(ii) scharakteryzować środkowoeuropejską różnorodność genetyczną bezpośrednio przedpojawieniem się CW,
(iii) określić datę, w której osoby o rodowodzie stepowym podobnym do Yamnaya pojawiły się po raz pierwszy w Europie Środkowej i zrozumieć ich pochodzenie genetyczne i strukturę społeczną,
(iv) scharakteryzować rodzaj i zakres wymiany biologicznej między „miejscowymi” i „migrantami”po pojawieniu się CW oraz
(v) zidentyfikować przekształcenia społeczne związane ze zmianami genetycznymi i archeologicznymi.
RESULTS
General sample overview
WYNIKIOgólny przegląd próbek
We screened 261 prehistoric individuals (table S3) from 37 sites (table S2) for ancient human DNA preservation, of which 219 individuals were enriched for 1,233,013 ancestry informative sites in the human genome (“1240k capture panel”) (8). After enrichment, individuals with fewer than 30,000 sites covered (on 1240k) or signs of contamination were removed (n = 13), resulting in a dataset of 206 newly reported individuals. We combined our dataset with 65 previously published individuals (5, 61, 62) from Bohemia (with >30,000 covered sites on 1240k; table S4) and wider (table S5), thereby extending the total number of published Bohemian Neolithic and pre-CW Eneolithic individuals from 7 to 58 (fig. S2), CW individuals from 7 to 54 (fig. S3), BB individuals from 40 to 64 (fig. S4), and EBA individuals from 11 to 95 (fig. S5). Crucially, we substantially expand the sample size of individuals around the time of CW formation (~3200 to 2600 BCE, from n = 1 to n = 50; Fig. 1B), i.e., the last pre-CW (Baden, Řivnáč, and GAC, from n = 0 to n = 18) and the early CW (from n = 1 to n = 32) individuals, allowing us to directly study the origin of CW in central Europe, the nature of their migration, and social interactions with coexisting pre-CW people. First-degree relatives were excluded from allele frequency–based analyses [f statistics, qpWave, qpAdm, Distribution of Ancestry Tracts of Evolutionary Signals (DATES), and Y chromosome analyses; table S4 and see Materials and Methods]. We also report 140 new radiocarbon dates to aid in finer temporal resolution, allowing us to study the genetic changes between early and late phases of important third millennium BCE cultural groups (e.g., CW, BB, and Únětice; tables S4 and S6).
Przebadaliśmy 261 prehistorycznych osobników (tabela S3) z 37 miejsc (tabela S2) pod kątem zachowania starożytnego ludzkiego DNA, z czego 219 osobników zostało wzbogaconych o 1, 233, 013 informacji o przodkach w ludzkim genomie („panel przechwytywania 1240k”) (8). Po wzbogaceniu usunięto osobniki z mniej niż 30 000 zakrytych miejsc (na 1240 tys.) lub oznaki skażenia (n = 13), co dało zestaw danych obejmujący 206 nowo zgłoszonych osobników. Połączyliśmy nasz zestaw danych z 65 wcześniej opublikowanymi osobami (5, 61, 62) z Czech (z ponad 30 000 objętych stanowiskami na 1240 tys.; tabela S4) i szerszych (tabela S5), rozszerzając w ten sposób całkowitą liczbę danych opublikowanych czeskiego neolitu i pre-CW Eneolityczne z 7 do 58 (ryc. S2), CW z 7 do 54 (ryc. S3), BB z 40 do 64 (ryc. S4) i EBA z 11 do 95 (ryc. S5). Co najważniejsze, znacznie zwiększamy wielkość próby osobników w czasie powstawania CW (~3200 do 2600 p.n.e., od n = 1 do n = 50; ryc. 1B), tj. ostatniego okresu przed CW (Baden, Řivnáč i GAC, od n = 0 do n = 18) i wczesne CW (od n = 1 do n = 32) osobników, co pozwala nam bezpośrednio zbadać pochodzenie CW w Europie Środkowej, charakter ich migracji i interakcje społeczne z współistniejącymi sprzed CW ludźmi . Krewni pierwszego stopnia zostali wykluczeni z analiz opartych na częstości alleli [statystyki f, qpWave, qpAdm, Distribution of Ancestry Tracts of Evolutionary Signals (DATES) i analizy chromosomu Y; tabela S4 i patrz Materiały i metody]. Zgłaszamy również 140 nowych datowań radiowęglowych, aby pomóc w dokładniejszej rozdzielczości czasowej, co pozwala nam badać zmiany genetyczne między wczesną i późną fazą ważnych grup kulturowych trzeciego tysiąclecia pne (np. CW, BB i Únětice; tabele S4 i S6).
Bohemia before Corded Ware (pre-CW, before ~2800 BCE)
We first assessed the genome-wide data by projecting the ancient individuals from Bohemia onto the first two axes of a principal components analysis (PCA) constructed from 1141 modern-day West Eurasian individuals (table S7). In the resulting PCA plot (Fig. 2A), all (n = 58) pre-CW individuals from Bohemia plot between Anatolia_Neolithic and Western HG (WHG), in close proximity to published culturally pre-CW individuals from central Europe (3, 8, 11, 13, 25). This suggests an absence of steppe ancestry, which we formally confirmed using qpAdm modeling (table S8), revealing that pre-CW individuals from Bohemia can be largely modeled as two-way mixtures of Anatolia_Neolithic and WHG (Fig. 3A, tables S8 and S9, and fig. S6). The percentage of HG ancestry is positively correlated with time (Spearman’s rank correlation r = 0.39, P < 0.004), showing that the previously reported trend of increasing HG ancestry during the Neolithic also took place in Bohemia (3, 11). We found this HG ancestry increase to be best modeled as a two-stage linear process (Fig. 3A, table S8, and the Supplementary Materials), with an increase in HG ancestry during the fifth millennium BCE, followed by stasis (nonsignificant slope) thereafter (Fig. 3A and Supplementary Materials).
Bohemia przed ceramiką sznurową (przed CW, przed ok. 2800 p.n.e.)
Najpierw oceniliśmy dane całego genomu, rzutując starożytne osobniki z Czech na pierwsze dwie osie analizy głównych składowych (PCA) skonstruowanej z 1141 współczesnych osobników z Zachodniej Eurazji (tabela S7). Na wynikowym wykresie PCA (ryc. 2A) wszystkie (n = 58) osobniki przed CW z obszaru Czech między Anatolia_Neolithic a zachodnim HG (WHG), w bliskiej odległości od opublikowanych osobników kulturowo przed CW z Europy Środkowej (3, 8 , 11, 13, 25). Sugeruje to brak przodków stepowych, co formalnie potwierdziliśmy za pomocą modelowania qpAdm (tabela S8), ujawniając, że osobniki sprzed CW z Czech można w dużej mierze modelować jako dwukierunkowe mieszaniny Anatolia_Neolithic i WHG (ryc. 3A, tabele S8 i S9 i rys. S6). Odsetek pochodzenia HG jest dodatnio skorelowany z czasem (korelacja rang Spearmana r = 0,39, P < 0,004), co pokazuje, że wcześniej zgłoszony trend wzrostu pochodzenia HG w okresie neolitu miał również miejsce w Czechach (3, 11). Odkryliśmy, że ten wzrost pochodzenia HG najlepiej modelować jako dwuetapowy proces liniowy (ryc. 3A, tabela S8 i materiały uzupełniające), ze wzrostem pochodzenia HG podczas piątego tysiąclecia p.n.e., po którym następuje staza (nieistotne odchylenie) następnie (ryc. 3A i materiały uzupełniające).
**** (skrót – pełny tekst u źródła)
Together, these results indicate a largely (significantly more than 50%) nonlocal genetic origin of Bohemian Funnelbeaker individuals.
The second such case can be seen in the Řivnáč to GAC cultural transition. GAC individuals carry the most HG ancestry among pre-CW cultural groups from Bohemia (25.7%, ±1.4), significantly more than Řivnáč individuals [f4(Mbuti.DG, WHG; Řivnáč, GAC) >> 0; z score, 4.46]. However, the estimated date of HG admixture in GAC is not later than in Řivnáč individuals (Fig. 3B and table S11), suggesting that GAC individuals do not descend from a recent mixture of Řivnáč and an HG source but instead constituted a recent, nonlocal incursion in Bohemia from a region that received more HG gene flow [e.g., Poland (13, 63)], in accordance with interpretations of archaeological evidence (56).
Łącznie wyniki te wskazują w dużej mierze (znacznie ponad 50%) na nielokalne pochodzenie genetyczne osobników z czeskiego kręgu kultury pucharów lejkowatych.
Drugi taki przypadek można zaobserwować w przejściu kulturowym z kultury Řivnáč do kultury GAC.Osobniki GAC mają największy udział pochodzenia od HG wśród grup kulturowych sprzed kultury CW z Czech (25,7%, ±1,4), znacznie więcej niż osoby z Řivnáč [f4(Mbuti.DG, WHG; Řivnáč, GAC) >> 0;z punktacją 4,46].Jednak szacowana data domieszki HG w GAC nie jest późniejsza niż u osobników Řivnáč (ryc. 3B i tabela S11), co sugeruje, że osobniki GAC nie pochodzą z niedawnej mieszaniny Řivnáč i źródła HG, ale ich źródło stanowią niedawne, nielokalnenajazdy w Czechach z regionu, który otrzymał większy przepływ genów HG [np. Polska (13, 63)], zgodnie z interpretacjami dowodów archeologicznych (56).
A distinct genetic origin for Řivnáč and GAC individuals is further supported by PCA and qpAdm modeling. From PCA, we find that with the exception of TUC003, Řivnáč and GAC individuals form distinct clouds (Fig. 3C). This is confirmed by qpAdm modeling where GAC individuals can be modeled as a mixture of Anatolia_Neolithic and Loschbour but not Anatolia_Neolithic and Körös_HG, while the opposite is true for Řivnáč individuals (table S14). Consequently, Řivnáč and GAC individuals are distinguishable based on the amount and source of HG ancestry, suggesting that Bohemia was inhabited by genetically differentiated groups of Řivnáč and GAC individuals at the time of CW appearance. The Řivnáč outlier (TUC003) also raises the interesting possibility of an individual born into a GAC but buried in a Řivnáč cultural context.
Among the 16 Řivnáč and GAC individuals who are contemporaneous with or postdate the appearance of CW in Bohemia (Fig. 1B), we find no detectable traces of steppe ancestry (Fig. 2A and table S8), suggesting that biological exchange from CW/Yamnaya into culturally pre-CW people (e.g., Řivnáč and GAC) was low, possibly nonexistent. Steppe ancestry coappears with CW individuals in early third millennium BCE Bohemia.
Odmienne pochodzenie genetyczne osobników Řivnáč i GAC jest dodatkowo wspierane przez modelowanie PCA i qpAdm. Z PCA dowiadujemy się, że z wyjątkiem TUC003, osobniki Řivnáč i GAC tworzą wyraźne chmury (ryc. 3C). Potwierdza to modelowanie qpAdm, w którym osobniki GAC można modelować jako mieszaninę Anatolia_Neolithic i Loschbour, ale nie Anatolia_Neolithic i Körös_HG, podczas gdy w przypadku osobników Řivnáč jest to odwrotne (tabela S14). W związku z tym osobniki Řivnáč i GAC można rozróżnić na podstawie liczby i źródła pochodzenia HG, co sugeruje, że w momencie pojawienia się CW Czechy były zamieszkiwane przez zróżnicowane genetycznie grupy osobników Řivnáč i GAC. Obserwacja odstająca Řivnáč (TUC003) również wskazuje na interesującą możliwość, że mamy osobę urodzoną w GAC, ale pochowaną w kulturowym kontekście Řivnáč. Wśród 16 osobników Řivnáč i GAC, którzy są współcześnie lub po pojawieniu się CW w Czechach (ryc. 1B), nie znajdujemy wykrywalnych śladów przodków stepowych (ryc. 2A i tabela S8), co sugeruje, że wymiana biologiczna z CW/Yamnaya na ludzi przed CW (np. Řivnáč i GAC) była niska, prawdopodobnie nieistniejąca. Rodowód stepowy występuje razem z osobnikami CW na początku trzeciego tysiąclecia p.n.e. w Czechach.
Corded Ware
We report genomic data from the earliest CW individuals to date, including STD003 (northwestern Bohemia, 3010 to 2889 calibrated (cal) BCE), VLI076 (central Bohemia, 3018 to 2901 cal BCE), OBR003 (central Bohemia, 2911 to 2875 cal BCE), and PNL001 (eastern Bohemia, 2914 to 2879 cal BCE), showing that CW was widespread across Bohemia by 2900 BCE. The early radiocarbon dates are also supported by these individuals’ genetic profiles, who occupy the most extreme positions on PC2 (Fig. 2B), as expected under a scenario of the earliest CW being migrants from the east who mixed with locals, resulting in intermediate PC2 positions in later generations.
Ceramika Sznurowa
Przedstawiamy dane genomowe od najwcześniejszych osobników CW do tej pory, w tym STD003 (północno-zachodnie Czechy, 3010 do 2889 kalibrowane (cal) BCE), VLI076 (środkowe Czechy, 3018 do 2901 cal BCE), OBR003 (środkowe Czechy, 2911 do 2875 cal BCE) i PNL001 (wschodnie Czechy, od 2914 do 2879 cal pne), pokazując, że CW było szeroko rozpowszechnione w Czechach do 2900 pne.Wczesne daty radiowęglowe są również poparte profilami genetycznymi tych osób, które zajmują najbardziej ekstremalne pozycje na PC2 (ryc. 2B), zgodnie z oczekiwaniami w scenariuszu, w którym najwcześniejsza CW to migranci ze wschodu, którzy mieszali się z miejscowymi, co skutkuje pośrednimiPozycjami PC2 w późniejszych generacjach.
To explore the formation of the Bohemian CW gene pool, we grouped CW individuals with steppe ancestry and mean age > 2600 BCE (n = 27) into a Bohemia_CW_Early group and the rest (n = 21) into Bohemia_CW_Late (table S4). We found poor statistical support (P < 0.005) for modeling Bohemia_CW_Early as a two-way mixture of any known Yamnaya source and any local Bohemian or nonlocal pre-CW source from Poland, Ukraine, Hungary, or Germany (table S15). When using distal sources as proxies for the Neolithic ancestry (Anatolia_Neolithic and a range of HG sources), we found no strong support (P < 0.05) for all but one of the three-way distal models (table S16). However, this one statistically supported model results in a previously unobserved ratio of Neolithic ancestry in Europe (i.e., a Neolithic population of ~1:1 ratio of Anatolia_Neolithic:Sweden_Motala_HG). In addition, when modeling early CW individually as “standard” three-way mixtures of Anatolia_Neolithic, WHG, and Yamnaya_Samara (3), we find that in 37% (10 of 27) of cases, the model lacks strong support (P < 0.05 or infeasible; fig. S6 and table S9).
Aby zbadać tworzenie puli genów Bohemian CW, pogrupowaliśmy osobniki CW o pochodzeniu stepowym i średnim wieku > 2600 pne (n = 27) w grupę Bohemia_CW_Early, a resztę (n = 21) w Bohemia_CW_Late (tabela S4). Znaleźliśmy słabe wsparcie statystyczne (P < 0,005) dla modelowania Bohemia_CW_Early jako dwukierunkowej mieszanki dowolnego znanego źródła Yamnaya i dowolnego lokalnego źródła czeskiego lub nielokalnego przed CW z Polski, Ukrainy, Węgier lub Niemiec (tabela S15). Używając źródeł dystalnych jako danych zastępczych dla przodków neolitu (Anatolia_Neolithic i szereg źródeł HG), nie znaleźliśmy silnego poparcia (P < 0,05) dla wszystkich modeli trójczynnikowych dystalnych poza jednym (tabela S16). Jednak ten jeden statystycznie potwierdzony model skutkuje wcześniej nieobserwowanym stosunkiem przodków neolitu w Europie (tj. populacja neolitu wynosi ~1:1 stosunek Anatolia_Neolithic:Szwecja_Motala_HG). Ponadto, podczas indywidualnego modelowania wczesnego CW jako „standardowych” trójczynnikowych mieszanek Anatolia_Neolithic, WHG i Yamnaya_Samara (3), stwierdzamy, że w 37% (10 z 27) przypadków modelowi brakuje silnego wsparcia (P < 0,05). lub jest ono niewykonalne; rys. S6 i tabela S9).
*** skrót
We provide the first genomic data from CW individuals without steppe ancestry, thereby elucidating the social processes of interaction between CW and pre-CW people. Observing only females (four of four) among early CW individuals without steppe ancestry (Figs. 2B and 3C) suggests that the process of assimilating pre-CW people into early CW society was female-biased. Two of these females (STD003 and VLI008) plot in close PCA space to GAC individuals from Bohemia and Poland (Fig. 3C). When grouped together, we find that STD003+VLI008 share more genetic affinity with Bohemian GAC than with Bohemian Řivnáč [f4(Mbuti.DG, STD003+VLI008; Bohemia-GAC, Bohemia-Řivnáč) < 0; z score, −2.32]. These two females are not genetically closer to Bohemian compared to Polish GAC individuals [f4(Mbuti.DG, STD003+VLI008; Bohemia-GAC, Poland-GAC) ~ 0; z = 0.5], meaning that a nonlocal, (north)eastern origin (e.g., Poland) cannot be ruled out. In addition, VLI009 and VLI079 fall outside of the sampled Bohemian Middle Eneolithic (Baden, Řivnáč, and GAC) genetic variation in PCA, carrying significantly more HG ancestry (Fig. 3C and table S22), suggesting that a large proportion (50%, or higher when including STD003/VLI008) of the genetically pre-CW females of the early CW society originated from outside Bohemia.
Dostarczamy pierwsze dane genomowe od osobników CW bez przodków stepowych, wyjaśniając w ten sposób społeczne procesy interakcji między CW a ludźmi przed CW. Obserwacja tylko kobiet (cztery z czterech) wśród wczesnych osobników CW bez przodków stepowych (Ryc. 2B i 3C) sugeruje, że proces asymilacji osób przed CW do wczesnego społeczeństwa CW był wzmożony w odniesieniu do kobiet. Dwie z tych kobiet (STD003 i VLI008) wysiewają w bliskiej przestrzeni PCA do osobników GAC z Czech i Polski (ryc. 3C). Po zgrupowaniu okazuje się, że STD003+VLI008 mają większe powinowactwo genetyczne z Bohemian GAC niż z Bohemian Řivnáč [f4(Mbuti.DG, STD003+VLI008; Bohemia-GAC, Bohemia-Řivnáč) < 0; punktacja z, −2.32]. Te dwie kobiety nie są genetycznie bliżej Bohemian w porównaniu z polskimi osobnikami GAC [f4(Mbuti.DG, STD003+VLI008; Bohemia-GAC, Poland-GAC) ~ 0; z = 0,5], co oznacza, że nie można wykluczyć nielokalnego (północno)wschodniego pochodzenia (np. Polska). Ponadto VLI009 i VLI079 nie mieszczą się w próbkowanej zmienności genetycznej czeskiego środkowego eneolitu (Baden, Řivnáč i GAC) w PCA, niosąc znacznie więcej przodków HG (ryc. 3C i tabela S22), co sugeruje, że duży odsetek (50%, lub wyższy, włączając STD003/VLI008) genetycznych pre-CW kobiet z wczesnego społeczeństwa CW, pochodził spoza Czech.
***
In addition to autosomal genetic changes through time, we observe a sharp reduction in Y-chromosomal diversity going from five different lineages in early CW to a dominant (single) lineage in late CW (Fig. 4A). We used forward simulations to explore the demographic scenarios that could account for the observed reduction in Y-chromosomal diversity. Performing 1 million simulations of a population with a starting frequency of R1a-M417(xZ645) centered around the observed starting frequency in Bohemia_CW_Early (3 of 11, 0.27), we assessed the plausibility of this lineage reaching the observed frequency in Bohemia_CW_Late (10 of 11, 0.91) in the time frame of 500 years under a model of a closed population and random mating (Materials and Methods). We reject the “neutral” hypothesis, i.e., that this change in frequency occurred by chance, given a wide range of plausible population sizes. Instead, our results suggest that R1a-M417(xZ645) was subject to a nonrandom increase in frequency, resulting in these males having 15.79% (4.12 to 44.42%) more surviving offspring per generation relative to males of other Y-haplogroups. We also find that this change in Y chromosome frequency is extreme compared to the changes in allele frequencies at fully covered autosomal 1240k sites (P < 0.0003) within the same males, suggesting a process that disproportionately affected Y-chromosomal compared to autosomal genetic diversity, ruling out a population bottleneck as the likely cause. Our results suggest that the Y-lineage diversity in early CW males was supplanted by a nonrandom process [selection, social structure, or influx of nonlocal R1a-M417(xZ645) lineages] that drove the collapse in Y-chromosomal diversity. A simultaneous decline of Y-chromosomal diversity dating to the Neolithic has been observed across most extant Y-haplogroups (64), possibly due to increased conflict between male-mediated patrilines (65). We view that changes in social structure (e.g., an isolated mating network with strictly exclusive social norms) could be an alternative cause but would be difficult to distinguish in the underlying model parameters.
Oprócz autosomalnych zmian genetycznych w czasie, obserwujemy gwałtowne zmniejszenie różnorodności chromosomu Y, przechodząc od pięciu różnych linii we wczesnej CW do dominującej (pojedynczej) linii w późnej CW (ryc. 4A). Wykorzystaliśmy symulacje postępowe, aby zbadać scenariusze demograficzne, które mogą wyjaśnić obserwowane zmniejszenie różnorodności chromosomu Y. Przeprowadzając 1 milion symulacji populacji z początkową częstością R1a-M417(xZ645) skupioną wokół obserwowanej początkowej częstości w Bohemia_CW_Early (3 z 11, 0,27), oceniliśmy prawdopodobieństwo, że ta linia osiągnie obserwowaną częstotliwość w Bohemia_CW_Late (10 z 11, 0,91) w okresie 500 lat w modelu zamkniętej populacji i losowego kojarzenia (Materiały i metody). Odrzucamy hipotezę „neutralną”, to znaczy, że ta zmiana częstości nastąpiła przypadkowo, biorąc pod uwagę szeroki zakres prawdopodobnych rozmiarów populacji. Zamiast tego nasze wyniki sugerują, że R1a-M417(xZ645) podlegał nielosowemu wzrostowi częstości, w wyniku czego samce miały o 15,79% (4,12 do 44,42%) więcej przeżywającego potomstwa na pokolenie w porównaniu z samcami z innych haplogrup Y. Odkryliśmy również, że ta zmiana w częstotliwości chromosomu Y jest ekstremalna w porównaniu do zmian w częstości alleli w całkowicie pokrytych autosomalnych miejscach 1240k (P < 0,0003) u tych samych samców, co sugeruje proces, który nieproporcjonalnie wpłynął na chromosom Y w porównaniu z autosomalną różnorodnością genetyczną, wykluczając wąskie gardło populacji jako prawdopodobną przyczynę. Nasze wyniki sugerują, że zróżnicowanie linii Y u wczesnych mężczyzn CW zostało wyparte przez nielosowy proces [wybór, struktura społeczna lub napływ nielokalnych linii R1a-M417(xZ645)], który doprowadził do załamania się różnorodności chromosomu Y.Jednoczesny spadek różnorodności chromosomów Y datowany na neolit zaobserwowano w większości istniejących haplogrup Y (64), prawdopodobnie z powodu zwiększonego konfliktu między patrylinami, w których pośredniczą mężczyźni (65). Uważamy, że zmiany w strukturze społecznej (np. wyizolowana sieć kojarzeń ze ściśle wykluczającymi się normami społecznymi) mogą być alternatywną przyczyną, ale byłyby trudne do rozróżnienia w podstawowych parametrach modelu.
***
Bell Beaker
The earliest BB individuals occupy a similar position in PCA as CW individuals (Fig. 4B and fig. S7), suggesting a degree of genetic continuity. To explore the genetic origin of early BB individuals (Bohemia_BB_Early; mean date, >2400 BCE; n = 3), we modeled them as a two-way mixture between preceding and contemporaneous cultural groups. We found support for a local origin, although nonlocal alternatives cannot be ruled out (table S28). However, our Bohemia_BB_Early group consists of only three (female) individuals and is therefore likely limited in representativeness and resolution to discern source populations.
We find that late BB individuals (Bohemia_BB_Late; mean date, ≤2400 BCE; n = 56) carry significantly more Middle Eneolithic–like ancestry compared to Bohemia_BB_Early (table S29). To explore this genetic shift, we modeled the ancestry of Bohemia_BB_Late as a two-way mixture of Bohemia_BB_Early and local Middle Eneolithic sources (table S30), finding support for an additional ~20% local Middle Eneolithic–like ancestry in late compared to early BB.
Puchary Dzwonkowate
Najwcześniejsze osobniki BB zajmują podobną pozycję w PCA jak osobniki CW (ryc. 4B i ryc. S7), co sugeruje pewien stopień ciągłości genetycznej. Aby zbadać pochodzenie genetyczne wczesnych osobników BB (Bohemia_BB_Early; średnia data, >2400 p.n.e.; n = 3), zamodelowaliśmy je jako dwukierunkową mieszankę między poprzedzającymi i współczesnymi grupami kulturowymi. Znaleźliśmy wsparcie dla lokalnego pochodzenia, chociaż nie można wykluczyć nielokalnych alternatyw (tabela S28). Jednak nasza grupa Bohemia_BB_Early składa się tylko z trzech (żeńskich) osobników i dlatego jest prawdopodobnie ograniczona pod względem reprezentatywności i rozdzielczości do rozróżniania populacji źródłowych. Odkryliśmy, że późne osobniki BB (Bohemia_BB_Późne; średnia data ≤2400 p.n.e.; n = 56) mają znacznie więcej przodków podobnych do środkowego eneolitu w porównaniu z Bohemia_BB_Early (tabela S29). Aby zbadać tę zmianę genetyczną, zamodelowaliśmy pochodzenie Bohemia_BB_Late jako dwukierunkową mieszankę źródeł Bohemia_BB_Wczesnych i lokalnych środkowych eneolitycznych (tabela S30), znajdując poparcie dla dodatkowego ~20% lokalnego przodka podobnego do środkowego eneolitu pod koniec w porównaniu do wczesnego BB .
We observe a closer phylogenetic relationship between the Y chromosome lineages found in early CW and BB than in either late CW or Yamnaya and BB. R1b-L151 is the most common Y-lineage among early CW males (6 of 11, 55%) and one branch ancestral to R1b-P312 (Fig. 4A), the dominant Y-lineage in BB (5). Although it is not possible to determine whether the P312 mutation(s) occurred in one of the early CW R1b-L151 males from Bohemia, we note that most Bohemian BB males are further derived at R1b-L2/S116 (R1b1a1a2b1), in contrast to BB males from England, several of whom are derived at R1b-L21(R1b1a1a2c1), showing that English and Bohemian BB males cannot be descendants of one another, but rather diversified in parallel. A scenario of R1b-P312 originating somewhere between Bohemia and England, possibly in the vicinity of the Rhine (66, 67), followed by an expansion northwest and east is compatible with our current understanding of the phylogeography of ancient R1b-L151–derived lineages.
Obserwujemy bliższy związek filogenetyczny między liniami chromosomu Y znalezionymi we wczesnym CW i BB niż w późnym CW lub Yamnaya i BB.R1b-L151 jest najczęstszą linią Y wśród wczesnych samców CW (6 z 11, 55%) i jedną z gałęzi przodków R1b-P312 (ryc. 4A), dominującej linii Y w BB (5).Chociaż nie jest możliwe ustalenie, czy mutacja (mutacje) P312 wystąpiła u jednego z wczesnych samców CW R1b-L151 z Czech, zauważamy, że większość czeskich samców BB pochodzi dalej z R1b-L2/S116 (R1b1a1a2b1), w przeciwieństwie do tegodo samców BB z Anglii, z których kilka pochodzi z R1b-L21(R1b1a1a2c1), co pokazuje, że samce BB angielskie i czeskie nie mogą być potomkami siebie nawzajem, ale raczej zróżnicowane równolegle.Scenariusz R1b-P312 pochodzący gdzieś między Czechami a Anglią, prawdopodobnie w pobliżu Renu (66, 67), po którym następuje ekspansja na północny zachód i wschód, jest zgodny z naszym obecnym rozumieniem filogeografii starożytnych linii wywodzących się z R1b-L151.
EBA—Únětice culture
The transition to the EBA in Bohemia is associated with a positive shift in the coordinates of PC2, relative to preceding late BBs (Fig. 4B, fig. S7, and table S31). Admixture f3 statistics are most negative when EHG (Eastern HG) or WSHG (West Siberian HG) are used as a second source in addition to the geographically and temporally proximal Bohemia_BB_Late (table S32), suggesting a northeastern contribution to Bohemia_Únětice_preClassical. To find a suitable proxy for a potential additional source population, we modeled Bohemia_Únětice_preClassical as a two-way mixture of local Bohemia_BB_Late and various sources more positive on PC2 (table S33). We reject mixture models involving Bohemia_BB_Late and Yamnaya (Samara, P = 5.3 × 10−10; Kalmykia, P = 5.8 × 10−10; Ukraine, P = 7.3 × 10−12; and Caucasus, P = 3.2 × 10−15) or Bohemia_BB_Late and CW (early, P = 1.1 × 10−4; late, P = 5.4 × 10−6). We fail to reject a two-way mixture model of 63.5% Bohemia_BB_Early and 36.5% Bohemia_BB_Late (P = 0.29), suggesting a large (63.5%) contribution from an early BB lineage, which was largely unsampled during the late BB phase (2400 to 2200 BCE), but represents a potential new lineage at the dawn of the Bronze Age. The Y-chromosomal data suggest an even larger turnover. A decrease of Y-lineage R1b-P312 from 100% (in late BB) to 20% (in preclassical Únětice) implies a minimum 80% influx of new Y-lineages at the onset of the EBA.
However, aware of the limited resolution of Bohemia_BB_Early (small sample size, low resolution, and large SEs), we explored alternative models for preclassical Únětice individuals. All model fits improve when Latvia_BA is included in the sources, resulting in two additional supported models (table S33). A three-way mixture of Bohemia_BB_Late, Bohemia_CW_Early, and Latvia_BA (P value of 0.086) not only supports a more conservative estimate of 47.7% population replacement but also accounts for the Y-chromosomal diversity found in preclassical Únětice, with R1b-P312 from Bohemia_BB_Late, R1b-U106 and I2 from Bohemia_CW_Early, and R1a-Z645 from Latvia_BA (Fig. 4A).
Although the geographic origin of this new ancestry cannot be precisely located, three observations offer clues. First, the Latvia_BA ancestry that improves all model fits (table S33) suggests an ultimate northeastern origin. Second, Y-haplogroup R1a-Z645 appears in Bohemia (and wider central Europe) for the first time at the beginning of the EBA, a lineage previously fixed in Baltic and common in Scandinavian CW males (23, 24), supporting a north/northeastern genetic contribution. Third, an Únětice genetic outlier (VLI051, male, Y-haplogroup R1a-Z645; table S34) resembles individuals from Bronze Age Latvia (Fig. 2D) (68), providing direct evidence for migrants from the northeast.
We also detect a genetic shift in the transition from preclassical to classical Únětice, reflected in the decrease in PC2 coordinates for Únětice individuals dated after ~2000 BCE (Fig. 4B and fig. S7) and confirmed using qpWave (table S35) and f4 statistics (table S36). Bohemia_Únětice_Classical can be modeled as a mixture of Bohemia_Únětice_preClassical and a local Eneolithic source (table S37). In contrast to the genetic shift between late BB and preclassical Únětice, the Y-lineage diversity remains similar throughout both Únětice phases, suggesting assimilation and subtler social changes.
EBA — kultura Úněticka
Przejście do EBA w Czechach wiąże się z dodatnim przesunięciem współrzędnych PC2 w stosunku do poprzedzających je późnych BBs (ryc. 4B, ryc. S7 i tabela S31). Statystyki domieszki f3 są najbardziej negatywne, gdy EHG (Eastern HG) lub WSHG (West Siberian HG) są używane jako drugie źródło oprócz geograficznie i czasowo proksymalnego Bohemia_BB_Late (tabela S32), co sugeruje północno-wschodni wkład w Bohemia_Únětice_preClassical. Aby znaleźć odpowiedni wskaźnik zastępczy dla potencjalnej dodatkowej populacji źródeł, wymodelowaliśmy Bohemia_Únětice_preClassical jako dwukierunkową mieszankę lokalnego Bohemia_BB_Late i różnych źródeł bardziej pozytywnych na PC2 (tabela S33).
Odrzucamy modele mieszane z udziałem Bohemia_BB_Late i Yamnaya (Samara, P = 5,3 × 10−10; Kałmucja, P = 5,8 × 10−10; Ukraina, P = 7,3 × 10−12; i Kaukaz, P = 3,2 × 10−15) lub Bohemia_BB_Late i CW (wczesny, P = 1,1 × 10-4; późny, P = 5,4 × 10-6). Nie odrzucamy dwukierunkowego modelu mieszanki 63,5% Bohemia_BB_Early i 36,5% Bohemia_BB_Late (P = 0,29), co sugeruje duży (63,5%) udział wczesnej linii BB, która była w dużej mierze niepróbkowana w późnej fazie BB (2400 do 2200 p.n.e.), ale reprezentuje potencjalny nowy rodowód na początku epoki brązu.
Dane z chromosomu Y sugerują jeszcze większy obrót. Spadek linii Y R1b-P312 ze 100% (w późnym BB) do 20% (w przedklasycznych Úněticach) oznacza co najmniej 80% napływ nowych linii Y na początku EBA. Jednak zdając sobie sprawę z ograniczonej rozdzielczości Bohemia_BB_Early (mała wielkość próbki, niska rozdzielczość i duże SE), zbadaliśmy alternatywne modele dla przedklasycznych osobników Únětice. Wszystkie dopasowania modeli poprawiają się, gdy w źródłach uwzględniono Latvia_BA, co skutkuje dwoma dodatkowymi obsługiwanymi modelami (tabela S33). Trójczynnikowa mieszanka Bohemia_BB_Late, Bohemia_CW_Early i Latvia_BA (wartość P 0,086) nie tylko wspiera bardziej konserwatywne szacunki zastąpienia populacji 47,7%, ale także odpowiada za różnorodność chromosomu Y znalezioną w przedklasycznych Úněticach, z R1b-P312 z Bohemia_BB_Late , R1b-U106 i I2 z Bohemia_CW_Early i R1a-Z645 z Latvia_BA (ryc. 4A). Chociaż geograficzne pochodzenie tego nowego przodka nie może być dokładnie zlokalizowane, trzy obserwacje dostarczają wskazówek. Po pierwsze, pochodzenie Latvia_BA, które poprawia wszystkie dopasowania modeli (tabela S33), sugeruje ostateczne pochodzenie północno-wschodnie. Po drugie, haplogrupa Y R1a-Z645 pojawia się po raz pierwszy w Czechach (i szerszej Europie Środkowej) na początku EBA, linii wcześniej ustalonej w Bałtyku i powszechnej u skandynawskich samców CW (23, 24), wspierającej północ/ północno-wschodni wkład genetyczny. Po trzecie, genetyczny odstający element Únětice (VLI051, samiec, haplogrupa Y R1a-Z645; tabela S34) przypomina osobniki z Łotwy z epoki brązu (ryc. 2D) (68), dostarczając bezpośrednich dowodów na migrantów z północnego wschodu. Wykrywamy również genetyczne przesunięcie w przejściu od przedklasycznego do klasycznego Únětice, odzwierciedlone w spadku współrzędnych PC2 dla osobników Únětic datowanych na ~2000 p.n.e. (ryc. 4B i ryc. S7) i potwierdzonych za pomocą statystyk qpWave (tabela S35) i f4 (tabela S36). Bohemia_Únětice_Classical można modelować jako mieszaninę Bohemia_Únětice_preClassical i lokalnego źródła eneolitycznego (tabela S37). W przeciwieństwie do genetycznego przesunięcia między późnym BB a przedklasycznymi Unětice, różnorodność linii Y pozostaje podobna w obu fazach Únětice, co sugeruje asymilację i subtelniejsze zmiany społeczne.
DISCUSSION
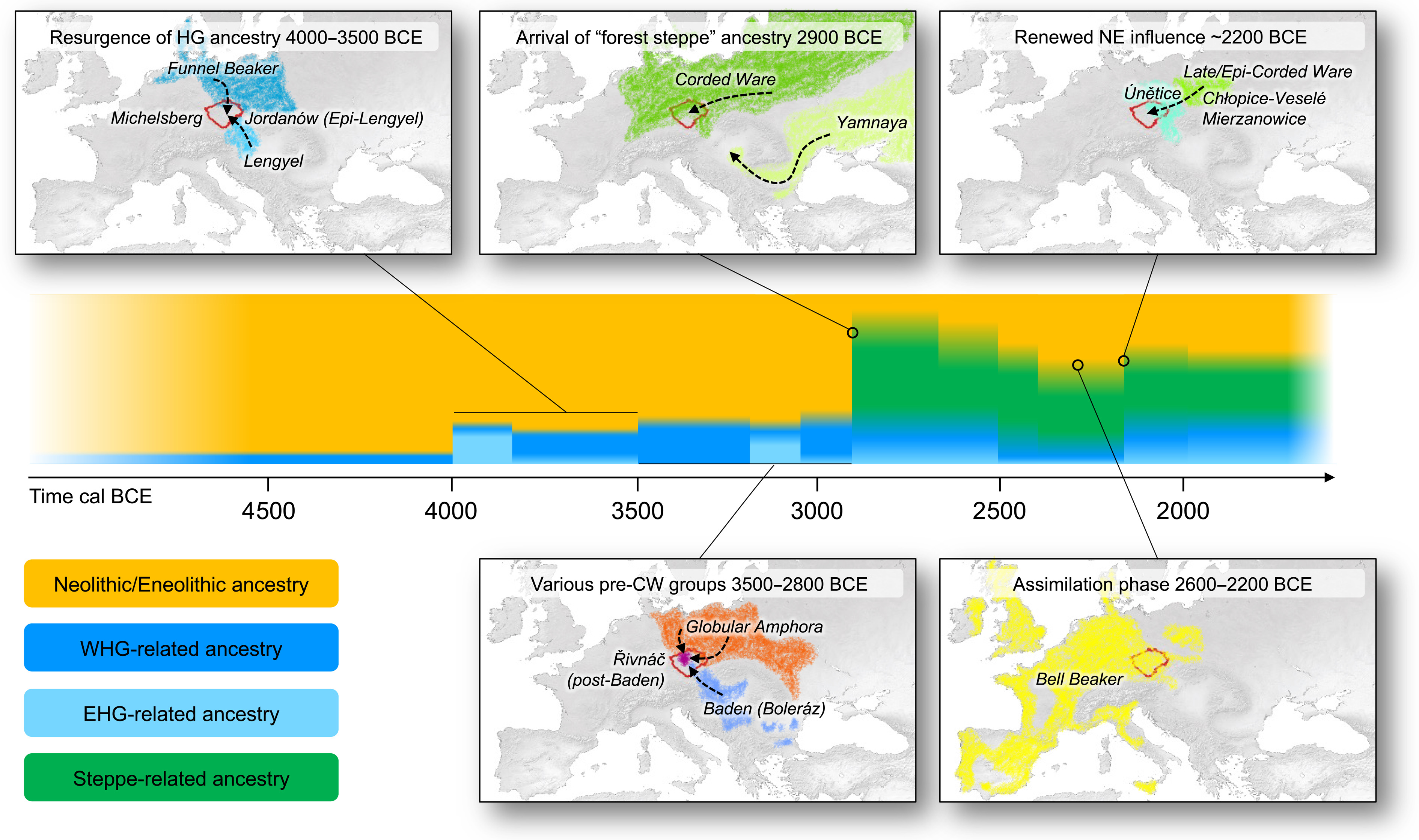
The high-resolution genetic time transect in Bohemia, allowing early and late phases of cultural groups to be divided and studied separately (e.g., CW, BB, and Únětice), elucidates several major processes before and after the arrival of steppe ancestry (Fig. 6). Our dense sampling allows detection of novel, important, and perhaps “unexpected” changes within cultural groups (e.g., CW and BB), if they are seen through a strict cultural-historical lens. Previous studies have largely been interpreted as revealing major migrations at the beginning and end of the Neolithic (i.e., periods where the incoming groups were genetically very distinct); however, our results reveal additional large genetic turnovers. By sampling consecutive and partially contemporaneous cultural groups, we show that the spread of Funnelbeaker and GAC (69, 70), as well as the origin of Únětice, involved large genetic shifts over short time periods, likely explained by migrations.
DYSKUSJA
Transekt czasu genetycznego o wysokiej rozdzielczości w Czechach, umożliwiający dzielenie i oddzielne badanie wczesnych i późnych faz grup kulturowych (np. CW, BB i Únětice), wyjaśnia kilka głównych procesów przed i po przybyciu przodków stepowych (ryc.6).Nasze gęste próbkowanie pozwala wykryć nowe, ważne i być może „nieoczekiwane” zmiany w obrębie grup kulturowych (np. CW i BB), jeśli są postrzegane przez ściśle kulturowo-historyczne soczewki.Wcześniejsze badania były w dużej mierze interpretowane jako ujawniające główne migracje na początku i na końcu neolitu (tj. okresy, w których przybywające grupy były bardzo odmienne genetycznie);jednak nasze wyniki ujawniają dodatkowe duże zmiany genetyczne.Próbując kolejne i częściowo współczesne grupy kulturowe, pokazujemy, że rozprzestrzenianie się pucharków lejkowatych i GAC (69, 70), a także pochodzenie Únětic, wiązały się z dużymi zmianami genetycznymi w krótkim okresie czasu, prawdopodobnie wyjaśnionymi migracjami.
fig.6. Schematyczne zestawienie głównych procesów, które kształtowały genetyczną i kulturową różnorodność Czech (czerwony kontur) na przestrzeni czasu.Strzałki na mapach wskazują raczej ogólny kierunek wpływów niż poszczególne trasy migracji.
We show that early CW were genetically exceptionally diverse, some resembling GAC and Yamnaya, with a few also falling outside of previously sampled central European Neolithic genetic diversity. Such a notably diverse signal is likely the result of the agglomeration of people from diverse cultural and linguistic backgrounds into an archaeologically similar but polyethnic or plural society. Important factors in ethnic identity include ancestry, history, ideology, and language (71, 72). The level of genetic differentiation (i.e., time since common ancestor) between early CW individuals with high and no steppe ancestry implies long biological isolation and hence different histories. The finding of GAC-like and Yamnaya-like genetic profiles in early CW suggests integration of people who came from ideologically diverse societies (i.e., neither GAC nor Yamnaya practiced strong gender differentiation in mortuary practices, unlike CW). It is likely that GAC and CW/Yamnaya individuals spoke different languages (3, 4, 43), meaning that early CW society in Bohemia encompassed people who had demonstrably different histories, likely originating from ideologically diverse cultures, who spoke different mother tongues.
Pokazujemy, że wczesne CW były wyjątkowo zróżnicowane genetycznie, niektóre przypominały GAC i Yamnaya, a kilka również nie mieściło się w wcześniej badanej różnorodności genetycznej neolitu środkowoeuropejskiego. Tak wyraźnie zróżnicowany sygnał jest prawdopodobnie wynikiem aglomeracji ludzi z różnych środowisk kulturowych i językowych w archeologicznie podobne, ale wieloetniczne lub pluralistyczne społeczeństwo. Ważne czynniki tożsamości etnicznej obejmują pochodzenie, historię, ideologię i język (71, 72). Poziom zróżnicowania genetycznego (tj. czas, jaki upłynął od wspólnego przodka) między wczesnymi osobnikami CW o wysokim pochodzeniu stepowym i bez pochodzenia stepowego implikuje długą izolację biologiczną, a zatem różne historie. Odkrycie profili genetycznych podobnych do GAC i podobnych do Yamnaya we wczesnym CW sugeruje integrację ludzi pochodzących ze społeczeństw zróżnicowanych ideologicznie (tj. ani GAC, ani Yamnaya nie praktykowały silnego zróżnicowania płci w praktykach pogrzebowych, w przeciwieństwie do CW). Jest prawdopodobne, że jednostki GAC i CW/Yamnaya mówiły różnymi językami (3, 4, 43), co oznacza, że wczesne społeczeństwo CW w Czechach obejmowało ludzi o wyraźnie odmiennej historii, prawdopodobnie pochodzących z różnych ideologicznie kultur, którzy mówili różnymi językami ojczystymi.
The assimilation process of individuals without steppe ancestry into early CW society was female-biased (43). However, finding females also among individuals with the highest amounts of steppe ancestry (3 of 5; Fig. 2B) suggests that they were also well represented among migrating CW individuals [in contrast to (43)] or perhaps assimilated from nearby Yamnaya groups (e.g., Hungary). Finding individuals without steppe ancestry in early CW contexts (n = 4) is more common than individuals with steppe ancestry in pre-CW contexts (e.g., GAC, n = 0). This pattern of asymmetric gene flow between the contemporaneous GAC and CW may reflect newcomers (CW groups) having more benefit from incorporating people with important local knowledge (i.e., from pre-CW cultural contexts) into their communities. The archaeological record shows continuity of such knowledge (e.g., pottery production and lithic raw materials) in several regions (22, 67, 73).
Vliněves is crucial for elucidating interactions between individuals with high and no steppe ancestry. This site yields the earliest dated CW (VLI076, 3018 to 2901 BCE) who is also genetically most differentiated from pre-CW individuals, while 20% (3 of 15) of the sampled early CW from Vliněves had no steppe ancestry. Intriguingly, we observe no archaeological differences between CW graves of individuals with and without steppe ancestry from two sites (Vliněves and Stadice; see the Supplementary Materials), suggesting full integration of genetically, and likely ethnically, diverse individuals within the same archaeological culture.
Finding Latvia_MN-like ancestry in early CW, in conjunction with the absence of Y-chromosomal sharing between early CW and Yamnaya males, suggests a limited or indirect role of known Yamnaya in the origin and spread of CW to central Europe. Our results allude to either a northeast European Eneolithic forest steppe contribution to early CW [a region consistent with some interpretations of the archaeological evidence (57)] or a hitherto unsampled steppe population who carried excess Latvia_MN-like ancestry, a scenario that is less likely given the high degree of genetic homogeneity among 3000-BCE steppe groups [e.g., Yamnaya and Afanasievo separated by ~2500 km but genetically almost indistinguishable (4, 61)]. As much of 4000- to 2500-BCE (north)eastern Europe remains unsampled, inferring the precise geographic origin of early CW individuals remains elusive.
Since social kinship systems influence patterns of genetic diversity (13, 42, 48, 74), it is likely that several different kin systems existed in third millennium BCE central Europe. The highly diverse genetic profiles (both nuclear and Y-chromosomal) of early CW suggest a different social organization to late CW and BB, whose Y chromosome pattern is indicative of strict patrilineality. This suggests that different cultural groups, in addition to using various forms of material culture and mortuary practices, likely also conformed to different ideologies as expressed in their mating pattern and/or social organization. This is supported by the finding of completely nonoverlapping Y chromosome variation between the partially contemporaneous late CW and BB, indicating a large degree of paternal mating isolation between these two groups, even when found at the same site (e.g., Vliněves).
The onset of the preclassical Únětice was accompanied by a ≥40% nuclear and ≥80% Y-chromosomal contribution ultimately originating from the northeast and breaking down the gender-differentiated mortuary practices and strict patrilineality of late CW and BB. This was neither evident in the burial customs nor in the material culture but could represent the underlying connection to the Baltics, the ultimate source of EBA amber in Bohemia associated with the later emerging Amber Road (75–77). Therefore, our results suggest two main periods (early CW and early Únětice) of genetic influence from the northeast, much of which remains unsampled in the European archaeogenetic record (e.g., Belarus).
Our results reveal a complex and highly dynamic history of Neolithic to EBA central Europe, during which migration and the movement of people facilitated abrupt genetic and social changes. Large-scale demic expansions occurred multiple times before and after the appearance of steppe ancestry in Europe. Early CW society was diverse and emerged amid a strong cultural and genetic transition, involving males and females of diverse origins and likely ethnicities. Genetic shifts occurred within CW, BB, and EBA societies despite continuity in material culture. Cultural affiliations played a major role in third millennium BCE social behaviors, which ultimately changed with the influx of new people over time. Although the impact of social processes is observable in patterns of genetic diversity, further interdisciplinary research is required to characterize the drivers of these changes, both at a micro- and macro-regional level.
Proces asymilacji jednostek bez przodków stepowych do wczesnego społeczeństwa CW był zorientowany na kobiety (43). Jednak znalezienie samic również wśród osobników o największym pochodzeniu stepowym (3 z 5; Ryc. 2B) sugeruje, że były one również dobrze reprezentowane wśród migrujących osobników CW [w przeciwieństwie do (43)] lub być może zasymilowane z pobliskich grup Yamnaya ( np. Węgry). Znajdowanie osobników bez pochodzenia stepowego we wczesnych kontekstach CW (n = 4) jest częstsze niż osobników o pochodzeniu stepowym w kontekstach sprzed CW (np. GAC, n = 0).
Ten wzorzec asymetrycznego przepływu genów między równoczesnym GAC i CW może odzwierciedlać nowoprzybyłych (grupy CW), które odnoszą większe korzyści z włączania ludzi o ważnej wiedzy lokalnej (tj. z kontekstów kulturowych sprzed CW) do swoich społeczności. Zapis archeologiczny wskazuje na ciągłość takiej wiedzy (np. produkcja ceramiki i surowców litowych) w kilku regionach (22, 67, 73). Vliněves ma kluczowe znaczenie dla wyjaśnienia interakcji między osobami o wysokim pochodzeniu stepowym i bez niego. Z tego miejsca pochodzi najwcześniej datowana CW (VLI076, 3018 do 2901 pne), która jest również najbardziej zróżnicowana genetycznie od osobników sprzed CW, podczas gdy 20% (3 z 15) pobranych wczesnych CW z Vliněves nie miało przodków stepowych.
Co ciekawe, nie obserwujemy żadnych różnic archeologicznych między grobami CW osób z i bez pochodzenia stepowego z dwóch stanowisk (Vliněves i Stadice; zob. Materiały Uzupełniające), co sugeruje pełną integrację genetycznie i prawdopodobnie etnicznie różnych osobników w ramach tej samej kultury archeologicznej. Odkrycie pochodzenia podobnego do Łotwy_MN we wczesnej CW, w połączeniu z brakiem podziału chromosomu Y między wczesnym CW i samcami Yamnaya, sugeruje ograniczoną lub pośrednią rolę znanej Yamnaya w powstawaniu i rozprzestrzenianiu CW w Europie Środkowej. Nasze wyniki nawiązują albo do wkładu stepów eneolitycznych Europy północno-wschodniej we wczesną CW [region zgodny z niektórymi interpretacjami dowodów archeologicznych (57)], albo do dotychczas niepróbkowanej populacji stepowej, która miała nadmierne pochodzenie przypominające Łotwę_MN, scenariusz, który jest mniej prawdopodobny biorąc pod uwagę wysoki stopień jednorodności genetycznej wśród grup stepowych 3000 pne [np. Yamnaya i Afanasievo rozdzielone około 2500 km, ale genetycznie prawie nie do odróżnienia (4, 61)].
Ponieważ większość z 4000 do 2500 pne (północno) wschodniej Europy pozostaje niepróbkowana, wnioskowanie o dokładnym pochodzeniu geograficznym wczesnych osobników CW pozostaje nieuchwytne. Ponieważ systemy pokrewieństwa społecznego wpływają na wzorce różnorodności genetycznej (13, 42, 48, 74), prawdopodobnie w trzecim tysiącleciu pne Europy Środkowej istniało kilka różnych systemów pokrewieństwa. Wysoce zróżnicowane profile genetyczne (zarówno autosomalny, jak i chromosom Y) wczesnego CW sugerują inną organizację społeczną niż późna CW i BB, których wzór chromosomu Y wskazuje na ścisłą patrylinearność. Sugeruje to, że różne grupy kulturowe, oprócz korzystania z różnych form kultury materialnej i praktyk pogrzebowych, prawdopodobnie również dostosowały się do różnych ideologii, co wyraża się w ich wzorcach godowych i/lub organizacji społecznej. Jest to poparte odkryciem całkowicie nienakładającej się zmienności chromosomu Y między częściowo równoczesnym późnym CW i BB, co wskazuje na duży stopień izolacji kojarzeń ojcowskich między tymi dwiema grupami, nawet jeśli występuje w tym samym miejscu (np. Vliněves).
Początkowi przedklasycznych Únětic towarzyszył ≥40% wkład autosomalny i ≥80% udział chromosomu Y, ostatecznie pochodzący z północnego wschodu i przełamujący zróżnicowane pod względem płci praktyki pogrzebowe oraz ścisłą patrylinearność późnych CW i BB. Nie było to widoczne ani w obyczajach pogrzebowych, ani w kulturze materialnej, ale mogło stanowić podstawę związku z krajami bałtyckimi, ostatecznym źródłem bursztynu EBA w Czechach, kojarzonym z powstającym później Szlakiem Bursztynowym (75–77). Dlatego nasze wyniki sugerują dwa główne okresy (wczesne CW i wczesne Únětice) wpływów genetycznych z północnego wschodu, z których większość pozostaje niepróbkowana w europejskim zapisie archeogenetycznym (np. Białoruś). Nasze wyniki ujawniają złożoną i bardzo dynamiczną historię od neolitu do EBA w Europie Środkowej, podczas której migracja i przemieszczanie się ludzi sprzyjało nagłym zmianom genetycznym i społecznym. Ekspansje demiczne na dużą skalę występowały wielokrotnie przed i po pojawieniu się przodków stepowych w Europie. Społeczeństwo wczesnego CW było zróżnicowane i wyłoniło się w trakcie silnej transformacji kulturowej i genetycznej, obejmującej mężczyzn i kobiety o różnym pochodzeniu i prawdopodobnym pochodzeniu etnicznym. Zmiany genetyczne wystąpiły w społeczeństwach CW, BB i EBA pomimo ciągłości w kulturze materialnej. Przynależności kulturowe odgrywały ważną rolę w zachowaniach społecznych trzeciego tysiąclecia p.n.e., które ostatecznie zmieniły się wraz z napływem nowych ludzi. Chociaż wpływ procesów społecznych można zaobserwować we wzorcach różnorodności genetycznej, konieczne są dalsze interdyscyplinarne badania, aby scharakteryzować czynniki napędzające te zmiany, zarówno na poziomie mikro-, jak i makroregionalnym.
Ta strona używa ciasteczek (cookies), w celach statystycznych. Możesz wyłączyć obsługę plików cookies w Twojej przeglądarce. Rozumiem
Privacy & Cookies Policy
Privacy Overview
This website uses cookies to improve your experience while you navigate through the website. Out of these, the cookies that are categorized as necessary are stored on your browser as they are essential for the working of basic functionalities of the website. We also use third-party cookies that help us analyze and understand how you use this website. These cookies will be stored in your browser only with your consent. You also have the option to opt-out of these cookies. But opting out of some of these cookies may affect your browsing experience.
Necessary cookies are absolutely essential for the website to function properly. This category only includes cookies that ensures basic functionalities and security features of the website. These cookies do not store any personal information.
Any cookies that may not be particularly necessary for the website to function and is used specifically to collect user personal data via analytics, ads, other embedded contents are termed as non-necessary cookies. It is mandatory to procure user consent prior to running these cookies on your website.