
Ksiądz Stanisław Pietrzak – Ród R1a macierzą Praindoeuropejczyków i Słowian
 Nowe wiadomości z genetyki rzutujące na genezę języków słowiańskich i indo-irańskich (R1a) oraz celtycko-germańskich (R1b) – warto przeczytać u księdza Stanisława Pietrzaka, tutaj tylko część nowego materiału, opublikowanego 22 marca 2016. CB:
Nowe wiadomości z genetyki rzutujące na genezę języków słowiańskich i indo-irańskich (R1a) oraz celtycko-germańskich (R1b) – warto przeczytać u księdza Stanisława Pietrzaka, tutaj tylko część nowego materiału, opublikowanego 22 marca 2016. CB:

Językowa wspólnota indo-europejska
Już blisko 250 lat temu uczeni stwierdzili istnienie jakiegoś podobieństwa i pokrewieństwa między językami Europy a językami Indii, a potem także Iranu, Pakistany i kilku tamtejszych krajów sąsiednich. Od tego czasu naukowcy pytają, czy i gdzie istniała jakaś ich mała praojczyzna, gdzie mogły powstać wspólne innowacje językowe i na jakich zasadach po rozpadzie pierwotnego języka praindoeuropejskiego powstały dialekty, już jako odrębne języki indoeuropejskie (grupy kentumowej i satemowej) i rozprzestrzeniły się na odległych sobie kontynentach: indyjskim i europejskim. Innymi słowy: jakie populacje przeniosły je z Europy na Wyżynę Irańską i Półwysep Indyjski lub – odwrotnie – stamtąd do Europy? Czy jest jakiś specjalny znak rozpoznawczy tej ludzkiej grupy, która by znalazła się po obydwu stronach: indo-irańskiej i europejskiej? Gdzie i kiedy ta praindoeuropejska grupa żyła i rozwijała się?
Niezmiernie wielką pomocą w rozwiązaniu tych pytań okazały się badania genealogii genetycznej i genetyki populacyjnej.
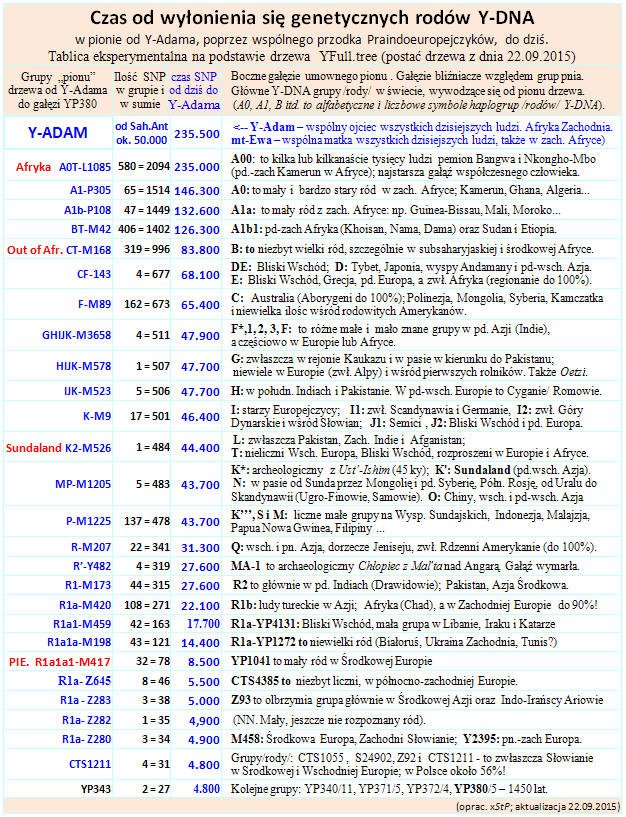
Dzięki genetyce, zwłaszcza genealogii Y-DNA już raczej wiemy, że tą ojczyzną, gdzie formowały się praindeoeuroejskie języki nie był jakiś geograficzny region, lecz migrujący w czasie od około 20.000 a.c. (przed Chr.) do około 6000 lat a.C. ze środkowo-wschodniej Azji do Środkowej Europy genetyczny ród R1a. Dzięki kolejnym, europejskim migracjom gałęzi tego rodu, uległy iondoeuropeizacji i bratni, zasadniczo zachodnioeuropejski ród R1b oraz staroeuropejskie rody I1 i I2, rozproszone prawie po całej Europie, zwłaszcza północnej, już w okresie polodowcowym, t.j. od około 12.000 lat a.C.
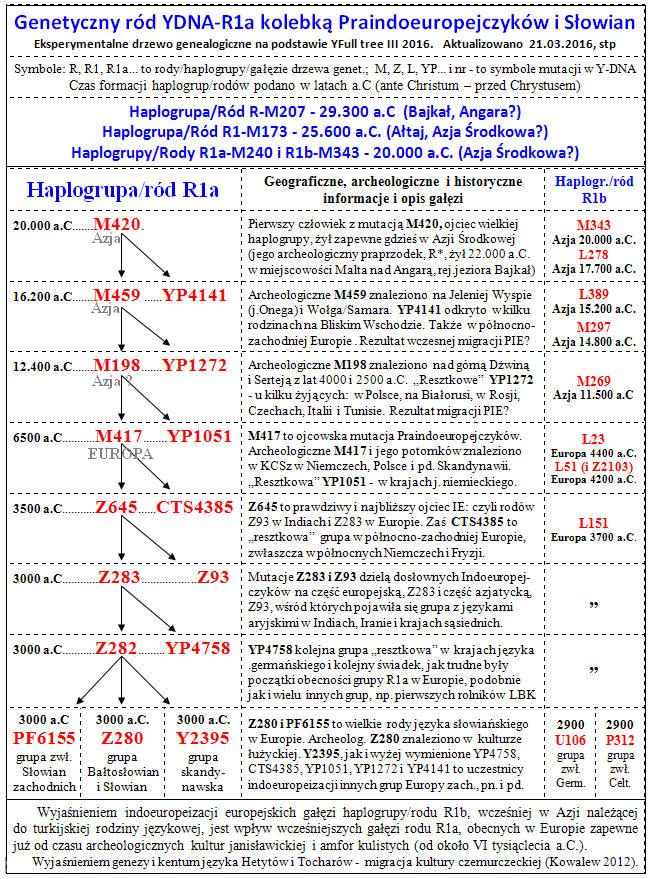
Rozwój rodu (genetycznej haplogrupy) R1a ilustruje poniższa tablica i drzewo genealogiczne Rodów R1a i (skrótowo) R1b
http://www.tropie.tarnow.opoka.org.pl/polacy1.htm#Praindoeuropejczycy
Słowianie, Prasłowianie, Praindoeuropejczycy. Slavs, Proto-Slavs, Proto-Indo-Europeans
| Ekstrakt Aktualizacja ekstraktu: 10.07.2015
Słowianie, Prasłowianie, Praindoeuropejczycy i ich pierwotna ojczyzna Podstawowe informacje genetyczne.
Poniżej na stronie przedstawiono wyniki rozmaitych analiz i badań językoznawczych, genetycznych i archeologicznych |
- Opis języków indoeuropejskich – IE
Odkryte w 1786 r. przez W. Jonesa wyraźne podobieństwo większości języków europejskich i głównych języków w Iranie i na Półwyspie Indyjskim skłoniło językoznawców do uznania ich pokrewieństwa i nadania im nazwy „języki indoeuropejskie” (nawiasem mówiąc bardziej logiczna i poprawna powinna być pisownia rozdzielna: języki indo-europejskie). Te związki językowe są świadectwem istnienia u ich podłoża jednego języka, wytworzonego i używanego przez jedną wspólnotę ludzką, żyjącą w zwartej grupie na stosunkowo niewielkim obszarze. Językowi temu nadano nazwę „praindoeuropejski”, a twórców i użytkowników tego języka, czyli współnotę praindoeuropejską nazwano „Praindoeuropejczykami” (pisownia w różnych krajach jest odmienna) – w skrócie: PIE.
Według dość powszechnie przyjętego w klasycznym językoznawstwie poglądu prawidło ewolucji języków, czyli glottochronologia, miałoby wskazywać na trzecie tysiąclecie przed Chr. jako na czas istnienia jeszcze jednego i w miarę niepodzielonego języka praindoeuropejskiego oraz jakiejś populacji Praindoeuropejczyków.
Gdyby chodziło o ojczyznę języka i wspólnoty praindoeuropejskiej to analiza najstarszego słownictwa, wspólnego dla wszystkich języków indoeuropejskich, miałaby wskazywać na stepy Europy Środkowej i południowej Ukrainy lub Rosji (choć nie brak uczonych wskazujących na Azję Środkową, Anatolię w Azji Mniejszej lub na Bałkany). Podział języka praindoeuropejskiego na znane nam dziś języki od krańców Europy po Półwysep Indyjski nastąpił na skutek rozprzestrzeniania się i migracji ludności oraz dokonywanych innowacji językowych w małych grupach, od siebie oddalonych oraz narażonych na kontakty (konwergencję) z zupełnie obcymi językami.
Cecha pierwotnego języka praindoeuropejskiego jest nadal dyskutowana, podobnie jak sam proces tzw. satemizacji i kentumizacji.
- Według jednych (Kluge, Reichert Sommerfelt, Kuryłowicz) w pierwotnym PIE istniały spółgłoski welarne i welarne spalatalizowane (zmiękczone k’, g’, gh’) – stan więc jakby przedsatemowy. Po rozpadzie wspólnoty w jednej jej części doszło do spirantyzacji owych spółgłosek welarnych palatalnych, czyli przeszły one w spółgłoski szczelinowe typu ś, sz, s. Na skutek tego np. słowo k’entum przybrało brzmienie satem (j. awestyjski) lub sto (j. słowiańskie) i stąd nazwa tej grupy języków: satemowa. W drugiej części spółgłoski welarne spalatalizowane utraciły miękkość. Na skutek tego np. słowo k’entum ‚sto’ przyjęło brzmienie kentum ‚sto’ i stąd nazwa tej grupy języków: kentumowa.
- Według drugich (Meillet, Hirt, Georgiev) w pierwotnym języku PIE był szereg spółgłosek welarnych (tylnojęzykowe k, g, gh) i welarnych labializowanych (wypowiadanych z ułożeniem warg ku „u”) – stan więc kentumowy, którego potwierdzeniem może być wczesna migracja kentumowego języka tocharskiego. Po rozpadzie wspólnoty w jednej części utrzymał się podział spółgłosek na welarne i labio-welarne; inne przemiany nie mają tu cech innowacji wspólnych. W drugiej części musiało dojść najpierw do palatalizacji pewnej części welarnych, a następnie do ich spirantyzacji, czyli satemizacji.
Proces satemizacji czy kentumizacji nie przechodził oczywiście bez zakłóceń z zewnątrz, zwłaszcza wewnętrznych niekonsekwencji w niektórych podgrupach, co dziś stwarza lingwistom ogromnie dużo kłopotów.
(Podstawowe informacje zob. Języki indoeuropejskie, t. 1 i 2, pod red. L. Bednarczuka, Warszawa 1986 oraz K. Polański [red]Encyklopedia językoznawstwa ogólnego, 1995).
Do niedawna uważano, że do językowej innowacji w postaci satemizacji doszło w centrum pierwotnego terytorium językowego; a ta innowacja nie dotarła do pozostającej w postaci kentumowej peryferii.
Jednak teraz językoznawstwo ma już doskonalsze narzędzie, umożliwiające dokładniejsze ustalenie warunków i okoliczności powstania wspólnoty języków indoeuropejskich oraz fundamentu podziału. Tym narzędziem jest genealogia Y-DNA. Ona bowiem wskazuje na istnienie konkretnej, biologicznej i pierwotnej wspólnoty praindoeuropejskiej w postaci populacji haplogrupy R1. Obecnie łatwo dostrzec, że językami kentumowymi wyróżniają się przede wszystkim populacje haplogrupy R1b (celtyckie, romańskie i germańskie).
Natomiast językami satemowymi wyróżniają się przede wszystkim populacje haplogrupy R1a (słowiańskie i indoirańskie). Śladem tego, że odmiana kentumowa była wcześniejszą przed satemową formą języka praindoeuropejskiego jest np. kentumowa postać starożytnego języka tocharskiego w Zachodnich Chinach. Badania genetyczne Li et al. 2010 archeologicznych szczątków siedmiu mężczyzn w Xiaohe (Xinjiang), identyfikowanych, choć niepewnie, z Tocharami, wykazały obecność tylko haplogrupy R1a1a1.
Można więc wnioskować, że do pierwotnego podziału języka w formie proto-praindoeuropejskiej na dwie podstawowe grupy doszło dopiero wtedy, gdy synowskie haplogrupy R1a i R1b rozeszły się demograficznie i terytorialnie. Ten podział populacji musiał nastąpić wnet po podziale haplogrupy R1 i powstaniu haplogrup R1a i R1b, około 10.000 lat przed Chr., skoro nie doszło do znacznego zmieszania się populacji co do Y-DNA. Ważny etap rozwoju językowej rodziny praindoeuropejskiej trzeba widzieć w Europie środkowej, około 2.500 lat przed Chr., po ponownym spotkaniu praindoeuropejskiej populacji R1a z grupami R1b, rozwijającymi się zapewne w kulturach unietyckiej i pucharów dzwonowatych.
Określenie więc „Indoeuropejczycy”(a raczej bardziej logicznie i ortograficznie w rozdzielnej pisowni: „Indo-Europejczycy”, jak ang. „Indo-Europeans”), wskazujące dotąd tylko na geograficzną rozległość pokrewnych języków i ich użytkowników, jest teraz uzasadnione także pierwotną jednością ich języka i wspólnym pochodzeniem biologicznym jego twórców.
| DZISIAJ zdecydowana większość użytkowników języków indoeuropejskich odznacza się mutacją Y-DNA M173, stanowiącą haplogrupę R1, oraz jej bliskich potomków. Jeden z nich, z mutacją M420 stanowi haplogrupę R1a, a drugi, z mutacją M343, stanowi haplogrupę R1b.
Choć te dwie ostatnie haplogrupy Y-DNA, które można nazwać patriarchalnymi (ojcowskimi) rodami, powstały i rozeszły się bardzo dawno, bo może nawet około 10 tysięcy lat przed Chr. i potem raczej rzadko się do siebie zbliżały, gdyż zasadniczo nie zmieszały sie ze sobą, to jednak w swoich głównych populacjach zachowały, jakby wbrew wróżbom glottochronologii, jakieś pokrewieństwo językowe, które można by nazwać językową wspólnotą proto-indoeuropejską. Rozumna i zgodna z Ockhama zasadą oszczędności jest przestroga, by źródeł dzisiejszych języków indoeuropejskich szukać raczej u ich bliższych i dalszych, aż do najdalszych przodków, którzy tworzyli i przekazywali potomnym swoje języki, a tylko dla pojedynczych i mniejszościowych odchyleń i odrębności, jak Baskowie czy Etruskowie, szukać innych przyczyn i okoliczności. Ponowne, ale już większe zbliżenie się owych dwóch gałęzi nastąpiło dopiero w Europie, prawdopodobnie na terenie przyszłej polskiej ziemi, o czym będzie tu mowa. |
Interesujące jest, że język siostrzanej względem R1 haplogrupy R2 w południowych Indiach, drawidyjski, został uznany za pokrewny językom indoeuropejskim w ramach tzw. grupy języków nostratyckich jeszcze wtedy, gdy nie znano powiązań genetycznych. Bardziej dogłębne badania porównawcze prajęzyków ludności haplogrupy R2 (w Azji Centralnej i w Indiach) mogłyby okazać sie pożyteczne w tej kwestii.
W każdym razie wraz z wczesnym oddaleniem się od siebie tych dwóch populacji, dryfowały także i ich języki, różnicując się wolniej wśród grupy wschodnioeuropejskiej R1a, a bardziej intensywnie wśród zachodnioeuropejskiej grupy R1b, co było uzależnione zwłaszcza od bardzo zróżnicowanych warunków życia i zapewne dłuższego przebywania wśród ludów turkijskich.
* * * * *
Zestawienie starożytnych Y-DNA rodu R1a, które dają nam wskazówkę na europejską ojczyznę rodu PIE
1. W Europie – ancientY-DNA grupy resztkowe spod R1a1 i R1a1a oraz wielka rodzina R1a1a1-M417, PIE
http://s27.postimg.org/z29u09qxv/R1a_Europe_dates.png
2. W Azji zidentyfikowano ancientY-DNA z mutacją synowską Z93 względem europejskich M417 i Z465
Oczekuje się archeologicznych znalezisk Y-DNA R1a-M420 i mutacji synowskich.
http://s23.postimg.org/nen0yig57/R1a_Asia_dates.png
3. Dla orientacji – dzisiejsze rozprzestrzenienie się i zagęszczenie rodu R1a
http://s11.postimg.org/8lih8edcj/1177epk.png
* * * * *
Powstały już tysiące prac naukowych, mających na celu rozwikłanie istotnej dla historii Europy i świata zagadki – źródła indoeuropejskiej wspólnoty językowej i pierwotnej jej ojczyzny. Obecnie takie prace nadal powstają.
Niedawno ukazała się głośna publikacja znawcy kultur praindoeuropejskich w Europie: Dawid W. Anthony, The Horse the Weel and Language. How bronze-age riders from the Eurasian steppes shaped the modern world, Oxford 2007. Z dostarczonego przez autora opisu i datowania kultur archeologicznych oraz korzystając z danych genealogii Y-DNA, można by naszkicować pewien mechanizm powstania i geograficzną przestrzeń języków praindoeuropejskich, ale niestety – raczej inną, niż ta, proponowana przez D.Anthony i prowizorycznie już akceptowana przez wielu uczonych.
Haak et al. 2015, Massive migration from the steppe was a source for Indo-European languages in Europe
http://www.nature.com/nature/journal/vaop/ncurrent/full/nature14317.html
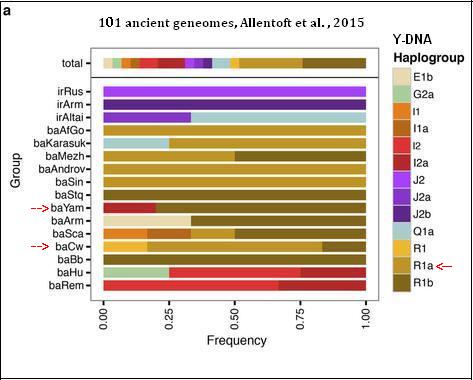
Badania Allentofta i zesp. 2015
| Chyba ostateczne rozwiązanie zagadki
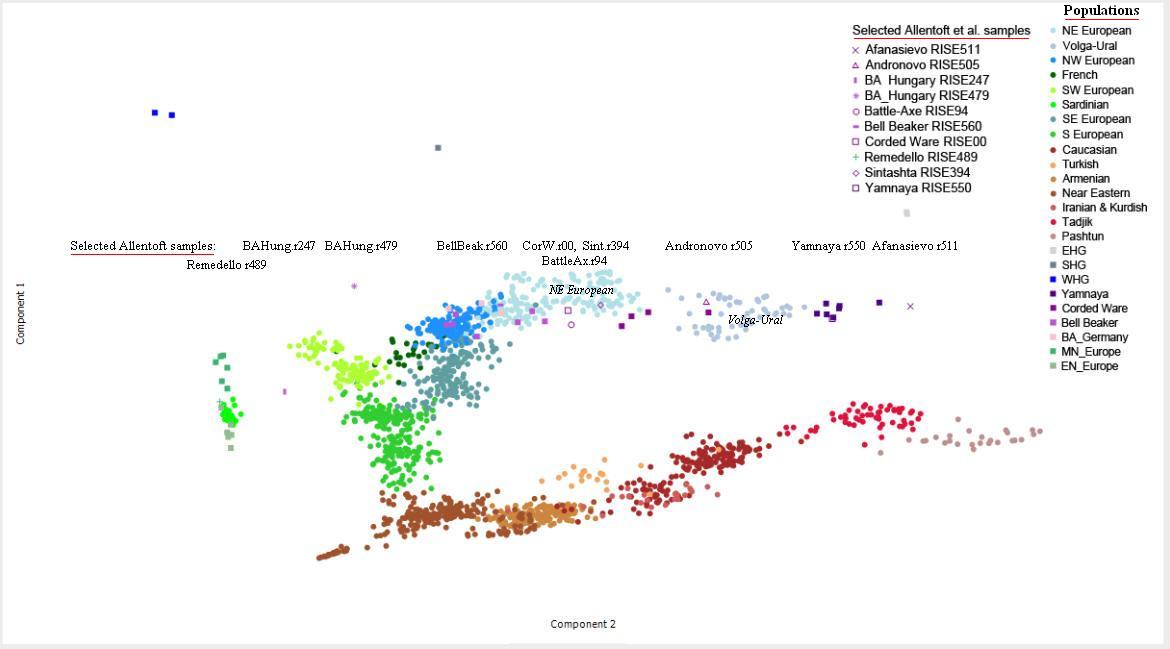
pochodzenia genetycznego rodu i ojczyzny Praindopeuropejczyków i Slowian. Objaśnienie pasków na diagramie irRus: epoka żelaza, pd. Rosja. Chociaż, zgodnie z najnowszą tradycją w nauce, najwięcej spodziewano się po archeologicznej kulturze jamowej (baYamna), że spełni ona wymogi bycia kolebką praindoeuropeizmu, to jednak okazało się, że w jej pięciu próbkach znaleziono jedną z przedindoeuropejskiej haplogrupy I2a (kol. ciemnoczerwony), a cztery – z R1b (kolor ciemnobrązowy) – podobnie jak w poprzednim badaniach Haak 2015, w których wszystkie dziesięć próbek z kultury jamowej okazały się z haplogrupy R1b. Ten fakt rzekomo upoważnił zespól Haaka, by areał kultury jamowej uznać za kolebkę praindoeuropeizmu. Nie znaleziono natomiast w tej kulturze, jak i poprzednio, ani jednej próbki właściwego praindoeuropejskiego rodu R1a, który – jako jedyny – łączy obydwa odległe rejony języka indoeuropejskiego: Europę i indo-irańskich Ariów w południowej Azji. Co więcej, genomy sąsiadującej z jamową kultury Sintaszta (Arkaim) i Andronowo w badaniach dystansu genetycznego okazały się najbliższe genomom z środkowoeuropejskiej kultury ceramiki sznurowej. W Sintaszta obydwie próbki wykazały mutację Z93, a wyróżnia ona migrantów z Europy do kultur stepowych w Azji oraz indo-irańskich Ariów; uwidacznia to kolor jasnobrązowy na powyższym diagramie. To odkrycia ma ogromne znaczenie historyczne, związane z genezą języków indoeuropejskich. Potwierdzeniem powyższych wyników jest zaprezentowana niżej genetyczna mapa odległości (PCA) autosomalnego DNA między poszczególnymi kulturami epoki brązu, a także w ich stosunku do ludności dzisiejszej Europy i Azji. |
{kind=link}
{kind=link}
{kind=link}
Aktualna (VII 2015) mapa rodu R1a w starożytnych grobach w Europie (w latach BP):
http://s4.postimg.org/lfkhruvn1/Datowanie_R1a.png
{kind=link}
……………………………………………………………………………………………………………..
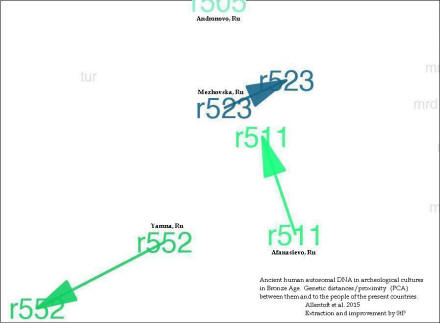
Z badań zespołu Allentoft et al. 2015 i innych grup
Niżej (dwa pliki jpg) – z powiększonego 12 krotnie obrazu mały wycinek mapy genetycznych odległości (PCA)
w autosomalnym DNA między ludnością kultur epoki brązu, a także między nimi a ludnością dzisiejszych krajów.

(Dla polepszenia czytelności można kliknąć w obydwie części obrazu, aby powiększyć).
Na ilustracji widzimy w skrócie nazwy dzisiejszych krajów, podane pismem koloru szarego. Zauważamy koncentrację DNA w rejonie kultury ceramiki sznurowej (r98 i r174) i polskiej unietyckiej (r150, kol. fioletowy) oraz Sintaszta i Andronowo (r395 i r505, kolor niebieski) w III tysiącleciu przed Chr. Widać ich bliskość do ludności dzisiejszej Polski i Chorwacji, oraz Węgier, Czech i Bułgarii (kolor szary). Widocznie noszą biologiczne dziedzictwo kultury ceramiki sznurowej i pokrewnych w Europie Środkowej (o polskim pochodzenie Chorwatów: http://www.doctrina.uph.edu.pl/doctrina%202013/1_doc_arkita.pdf ). Zauważamy także stosunkowo duży dystans do DNA kultury jamowej (Wołga-Ural, r552) i jej najbliższej krewnej – kultury Afanasjewo w południowej Syberii.
Inna forma prezentacji tego samego zjawiska
bliskości aDNA kultur Ceramiki Sznurowej, Unietyce i Sintaszta oraz dalekiej odleglości k. Jamowej i afansjewo
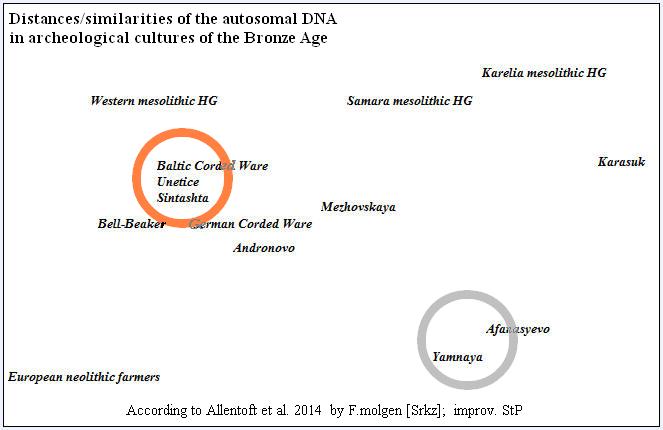
Jeszcze z innego programu ilustracja odległości PCA (otwórz) wybranych przedstawicieli starożytnych kultur archeologicznych
(101 gnomów od Allentoft et al. 2015). W kolumnie pierwszej – segmenty aDNA wybranych przedstawicieli kultur archeologicznych
z pracy Allentofta. W kolumnie drugiej uśrednione segmenty aDNA innych kultur (także ustalonych wirtualnie) od mezolitu
do epoki żelaza różnych regionów Europy i Azji dla porównania odległości. genetycznych.
{kind=link}
Można mieć teraz nadzieję, że kolejne, podobne badania archeogenetyczne ubogacą te wyniki, a nadto wskażą wcześniejszy etap życia rodu R1a, który wstępnie dostrzegam w kulturze amfor kulistych (związanej z Kujawami) i w kulturze janisławickiej (na przestrzeni od środkowego Dniepru na wschodzie do Kujaw na zachodzie). Sygnałem dla tej nadziei jest przybycie na Kujawy tak zwanych wkładek typu „Dęby” na początku VI tysiąclecia przed Chr. (L.Domańska 1990) oraz dwa sygnały obecności w bliskich Polsce rejonach gałęzi R1a z wczesną mutacją M459, np. datowane na czas około 5300 przed Chr. archeologiczne M459* na Jeleniej Wyspie (Jezioro Onega), wcześnie wydzielona spod M459 gałąź YP1272 z Wołynia oraz druga spod mutacji M459 archeologiczna M198* z około 4000 r. przed Chr. znad Górnej Dźwiny.
Badania: Czekunowa 2014, Haak 2015, Wencel 2015 i Nikitin 2012, Allentoft…
| Czas przełomu w badaniach i nadziei
W ostatnich kilku miesiącach opublikowano kilka istotnych dla kwestii genezy Praindoeuropejczyków i Prasłowian prac naukowych, które zaczynają stabilizować nasze poglądy na temat początku Praindoeuropejjczyków w Europie 1) Chekunova et al. 2014, The first results of genetic typing of local population and ancient humans in Upper Dvina region [ros.], https://www.academia.edu/9452168/Archaeology_of_lake_settlement_IV-II_mill._BC_Mazurkevich_A._Polkovnikova_M._Dolbunova_E._ed Mатериалы международной конференции, Посвященной Полувековому исследованию свайных Поселенийна северо-заПаде россии.санкт-Петербур, 13–15 ноября 2014 Г. 2) Haak et al. 2015, Massive migration from the steppe is a source for Indo-European languages in Europe 4) M.M. Wencel, 2015, An absolute chronological framework for the central-eastern European eneolithic, http://onlinelibrary.wiley.com/doi/10.1111/ojoa.12047/pdf
Pierwsza z nich informuje o odkryciu w archeologicznych szczątkach istotnej dla Praindoeuropejczyków i Prasłowian hapogrupy R1a-M198 ze stanowisk w kulturze Uświaty (Usvyaty) w osadzie Serteja (Serteya) nad górną Dźwiną Zachodnią, z datowaniami około 4000 i 2500 lat przed Chr. oraz nad pobliskim jeziorem Żyżyńskim, z datowaniami na około 2500 przed Chr. i 800-500 przed Chr. ……………………………………………………………………… ……………………………………………………………………………………….. To wszystko upoważnia mnie do powrotu do mojej koncepcji sprzed lat (dyskutowanej na forum Histmagu, DNA-Forums i w korespondencji z Jean Manco) że: Początku języka praindoeuropejskiego można upatrywać już w rodzie R1, gdzieś w Azji Środkowej. Tam w archeogenetyce ujawniła się haplogrupa R* (24.000 cal przed Chr.; Mal’ta nad Angarą pod Bajkałem). Na wschód od Bajkału, zwłaszcza w dorzeczu Amuru odkryto wczesne mutacje mitochondrialnej grupy C (C1a), która zapewne towarzyszyła migracji R1a-M420, a potem także R1a1-M459 ku Europie przez Kaukaz nad Dniepr (zob. wyżej) i dalej na Wołyń i Jelenią Wyspę na północy.
;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;; ;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;;
Na tle powyższych danych odczuwa się w naszym temacie pilną potrzebę wzmożenia prac archeologicznych i genetycznych nad: 2) kulturą janisławicko-rudoostrowską: http://pl.wikipedia.org/wiki/Kultura_janis%C5%82awicka 3) jej sąsiednią od północy – kulturą Narva: W nich należy szukać rozwoju społeczności i języka praindoeuropejskiego, a w słynnym „wschodnim exodusie” kultury amfor kulistych – już rozwiniętej ekspansji Praindoeuropejskiej społeczności i dialektów PIE (jak to podaję w poniższym artykule, po dokonaniu kilku poprawek). Natomiast migracje subkladów haplogrupy R1b1a, najpierw na wschód i południe, dokonała się na tereny jakieś języka prototurkijskiego. Ujawniona przez Haak i zesp. 2015 migracja na Zachód musiała dokonać się przez tereny satemowych wspólnot językowych rodu R1a, a także terutoria grup języka haplogrupy I1 i I2 (protogremańskiego?), co musiało owocować wzajemnymi przejęciami leksykalnymi czy gramatycznymi.
|
Teoria janisławicko-kujawska (extract)
Geneza wspólnoty praindoeuropejskiej
(ród, ojczyzna i język PIE).
( http://www.tropie.tarnow.opoka.org.pl/polacy1.htm#Praindoeuropejczycy )
Chociaż w nauce odnośnie lokalizacji ojczyzny językowej wspólnoty praindoeuropejskiej dość powszechnie akceptuje się teorię kurhanową, według której Praindoeuropejczycy formowali swoją społecznośc i język na terenie pomiędzy rzekami Donem i Uralem, to jednak wzbudza ona wiele zastrzeżeń i nie brak argumenów na rzecz wielu innych lokalizacji. Rozwijająca się od niedawna genealogia genomowa podpowiada jeszcze inną teorię i chyba ostateczne rozwiązanie tego problemu.
1. Praindoeuropejski ród R1a1. To, co jednoznacznie łączy ludzi podstawowych grup języka indoeuropejskiego w Europie i Azji, jest gentyczny ród z ojcowską mutacją M459, aktualnie oznaczany na drzewie współczesnego człowieka jako gałąź (haplogrupa) R1a1. Wyróżnia ona w Europie zwłaszcza ludy słowiańskie (do ok. 50% ludności; a w Polsce nawet 57%), a w Azji zwłaszcza indo-irańskich Ariów (lokalnie, jak nad Indusem w Pakistanie, do ok. 50% ludności, a w najwyższej kaście hinduskich braminów nawet 72%). Początek ekspansji rodu R1a1 datowany jest obecnie na około 5500 lat przed Chr. W linii prostej jest on potomkiem powstałej około 29.000 lat temu, może gdzieś w południowo-wschodniej Azji (Sundaland?) lub Azji środkowej, mutacji R1a-M420. Jakieś niesprzyjające warunki przez wiele tysiącleci tworzyly dla rozwoju tego rodu wąskie gardło, przez które zwycięsko przeszedł dopiero w Europie. Niewielkie odgałęzienia tego rodu R1a mogą znajdować się gdzieś w rejonie Płaskowyżu Irańskiego lub na Bliskim Wschodzie.
- W kręgu janisławicko-rodoostrowskim na Kujawach. Około połowy szóstego tysiąclecia przed Chr. niewielka grupa proto-indoeuropejskiej ludności, potomków tego właśnie ojcowskiego rodu R1a1-M459, migrując przez Bliski Wschód, gdzie w Anatolii zostawia jakąś część i ślady jej prajęzyka IE, przechodzi przez Kaukaz, ziemie nadczarnomorskie, Rudoj Ostrow nad Dnieprem i polskie Janisławice i przybywa na Kujawy. W archeologicznym kręgu janisławicko-rudoostrowskim, reprezentowanym szczególnie przez osadę Dęby 29 nad rzeką Bachorzą na Kujawach (stąd nazwa „teoria kujawska) ludność ta, jako jedyna, w pełni realizuje niesiony przez siebie model protoneolitycznej kultury kaukasko-nadczarnomorskiej. W sztuce wytwarzania narzędzi, używania materiałów krzemiennych i protoagrarnych tendencji odznacza się ona wyraźnymi cechami dystynktywnymi względem innych lokalnych kultur mezolitycznych, świadcząc równocześnie o swym odrębnym pochodzeniu i odrębności językowej.
Zapewne z tego kręgu kulturowego mała grupa dokonała migracji (łowieckiej?) w kierunku jeziora Onega, gdzie na Jeleniej Wyspie w archeologicznych szczątkach rozpoznano mutację R1a1-M459. Kolejno w rodzie tym pojawia się gałąź z mutacją YP1272, a zwłaszcza pokolenie M198, które oznaczone jest symbolem R1a1a. - W kulturze amfor kulistych. Po etapie rozwoju następuje zanik kultury Janisławickiej na Kujawach, spowodowany zapewne kryzysem demograficznym ludności haplogrupy R1a1. Ale około 4000 lat przed Chr. powstaje tu stopniowo jakby zupełnie nowa kultura, amfor kulistych, znów z wyraźnymi cechami dystynktywnymi. W nim genetyka domyśla się już rodu R1a1a1, z mutacją M417. Ta bowiem grupa rozpoznana została w dwóch grobach, przypisanych do zastępnej kultury, kultury ceramiki sznurowej. Wnet potem należu tu domyślać się mutacji Z645, której potomkowie rozdzielili się na dwa genetyczne rody: Z283 (europejska) i Z93 (na ogół azjatycka).
- Migracje ludności i dialektów IE. Około 3250 lat przed Chr. kultura amfor kulistych i jej ludność dociera już do środkowej Łaby i Solawy, gdzie zostaje objęta horyzontem kultur ceramiki sznurowej i gdzie w grobie z około 2400 roku zidentyfikowano haplogrupę R1a-M417 (Haak et al. 2015), oraz wcześniej tę samą mutację, ale w podgałęzi CTS4385, migrującą na zachód i północ Europy.
Nadto kultura amfor kulistych pojawia się około 2950 roku przed Chr. na Wołyniu, Podolu i Mołdawii, dochodząc nawet do środkowego Dniepru, a także na północno-wschodnich terenach, w dorzeczu Narwi, przy czym niektóre jej elementy docierają nawet do doliny Wołgi i podnóży Kaukazu. Jest to migracja rodu R1a1 i jego podgałęzi, zwłaszcza z mutacją “bałtosłowiańską” Z280 i Z92oraz “aryjską” Z93, a wraz z tą ludnością – migracja kultury i dialektów języka praindoeuropejskiego.
Obecne niektórych rosyjskich archeologów, antropologów i etnologów w pełni potwierdzają środkowo-europejskie pochodzenie praindoeuropejskich zabytków z kultur pucharów lejkowatcyh i amfor kulistych nad Dolnym Dnieprem, Donem i Wołgą, czyli w tzw. kulturze jamowej, oraz na północnym Kaukazie; a zarazem zaprzeczając teorii kurchanowej. - Indoeuropeizacja północnoeuropejskich gałęzi rodu i1 i i2. Terytorium kultur janisławicko-rudoostrowskiej i amfor kulistych prawie dosłownie nakłada się na terytorium wcześniejszxych tu kultur mezolitycznych – komornickiej i chojnicko-pieńkowskiej (będących we wspólnocie genetycznej i archeologicznej z ówczesną ludnością północnych Niemiec, Jutlandii i południowej Skandynawii, językowo zapewne pre-protogermańskiej) oraz neolitycznej kultury pucharów lejkowatych. Kultury te były tworzone zapewne przez staroeuropejskie rody (haplogrupy) I1 i I2 i neolitycznych migrantów znad Dunaju w kulturze późnolinearnej (z haplogrupami F*, G2a2b, I1-I2), To spotkanie musiało wpłynąć na wczesną ich indoeuropeizację, już około roku 4000 przed Chr.
6. Indoeuropeizacja ludności R1b. Przy spotkaniach ludności R1a w kulturze amfor kulistych i horyzontu sznurowego z bratnimi grupami italo-celtyckiej ludności R1b w archeologicznej kulturze pucharów dzwonowatych i unietyckiej (Wielkopolska, Śląsk, Czechy) doszło także do ich pełnej indoeuropeizacji językowej i powstania dialektów kentumowych, głównie zachodnioeuropejskich. Nie można jednak wykluczyć, że indoeuropejskie języki w europejskich gałęziach haplogrupie R1b tkwią jakoś już w haplogrupie R1, ojcowskiej dla R1a i R1b.
(Refleksja. Archeolodzy zauważają zaskakujący „fenomen miejsca” w pradolinie rzeki Bachorza na Kujawach, z miejscowościami zwłaszcza: Dęby, Dobre i Smarglin. Na tym niewielkim obszarze zarejestrowano „niezwykłe bogactwo różnorodnych tradycji kulturowych”, począwszy od protoneolitu, przez neolit, późny neolit, interstadium neolitu-brązu, do brązu w kulturze łużyckiej włącznie. Zidentyfikowano tu kilkanaście znanych w Polsce kultur lub grup archeologicznych z tamtych epok, przy czym wiele z nich zdradza na tle innych regionów swoją wczesność, podpowiadając, jakoby miejscowa ludność nad Bachorzą była ich inicjatorką albo przynajmniej niezwykle aktywną w przejmowaniu z bliższa lub dalsza cudzych inicjatyw kulturowych – i to do tego stopnia, że niekiedy trudno wskazać właściwego ich inicjatora i twórcę; zob. Czebreszuk, Makarowicz 1990; Czebreszuk, Ignaszak, Łoś 1997).
20.12.2014 (13.02.2015), StP.
:::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: ::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
( Janisławice-Kuyavian Theory, extract )
Proto-Indo-Europeans
The origin of the proto-Indo-European community
(family, homeland and PIE language)
http://www.tropie.tarnow.opoka.org.pl/polacy1.htm#Proto-Indo-Europeans
- The Proto-Indo-European family R1a. That’s what clearly brings basic group of people of Indo-European language in Europe and Asia, is a genetic family R1a. He distinguishes in Europe especially the Slavic peoples (up to approx. 50% of the population; in Poland 57%), in Asia especially Indo-Iranian Aryans (to approx. 50% of the population on Indus basin and to 72% among Bramins). It is now dated at about 5000 years before Christ. In the straight line the family is a descendant from mutation M420, created somewhere in south-east Asia (Sundaland?), which on the phylogenetic tree of modern man, marks the beginning of the great paternal branch R1a-M420, dated back to about 29000 years BP.
- In the archeological circle Janisławice-RudojOstrow on Kuyavia. In about half of the sixth millennium BC, a small group of the Proto-Indo-European population of this particular genetic paternal lineage R1a1-M459, migrating through the Middle East, where in Anatolia leaves some its part and traces its proto-IE language, passes through the Caucasus, lands on the Black Sea, Rudoj Ostrow on the Dnieper and Polish Janisławice and arrives at Kuyavia (Polish: Kujawy). This fact is supported by the R1a1*-M459 Oleni Ostrov in Karelia and R1a1b-YP1272 in Poland.
In the archaeological circle Janisławice-RudojOstrow, represented especially by the settlement Dęby 29 on the river Bachorza in Kuyavia, this population, as the only, fully implements their own model of the Caucasian-Black Sean proto-neolitic culture. In the craft of manufacturing tools, using flint and proto-agrocultural trend this culture is characterized by unique and distinct features, unseen in other local Mesolithic cultures, thus giving a proof to their separate origins and distinct language, probably already Indo-European. - In the Globular Amphora culture. After the development phase, Janisławice-RudojOstrov culture disappears, probably because of the demographic crisis of the population of the genetic haplogroup R1a1. About 4000 years before Christ, as if a completely new culture, Globular Amphora, arises gradually here, again with distinct features, which later geneticists guesses or recognize the haplogroup R1a1a-M198 and R1a1a1-M417 that distinguishes Proto-Indo-Europeans and their first migrations.
- 4. The migrations of the population and IE dialects. Around 3250 years BC the Globular Amphora culture and its population already reaches the middle Elbe and Saale rivers, where it is a subject to the Corded Ware horizon cultures and where geneticists have identified the haplogroup R1a-M417 (Haak et al. 2015) and probably in the sub-branches with mutations CTS4385 or Y2395, migrating to the west and north of Europe (Haak et al. 2008).
Moreover, about 2950 year BC the Globular Amphora culture appears in Volhynia, Podolia and Moldavia, and on the north-eastern from Poland on the Narew river basin (though some elements of the culture reaches up into the valley of the Volga and the foothills of the Caucasus). Is a migration of family Z645 and its subbranches Z283 and Z93, especially with the mutations „Balto-Slavic” Z280 and „Aryan” Z93, and with the population – migration of subsequent dialects of the proto-IE language. - Indo-Europeanization of the family I1 and I2. The territory of the Janisławice-RudojOstrow culture and Globular Amphora almost literally applies to the territory of the Mesolithic cultures: Komornicka and Chojnice-Pieńkowska and Neolithic Funnel Beaker Culture, probably created by the old-European families I1 and I2, which is in the genetical and archaeological community with the then population of northern Germany, Jutland and south Scandinavia, linguistically probably pre-proto-Germanic. This meeting must result in their early Indo-Europeanization.
- Indo-Europeanization of the family R1b. At meetings of the population R1a in the Globular Amphora culture and the Corded Ware horizon with the kindred groups of the populations of the Proto-Celtic R1b in the Bell Beaker and Unietice archaeological cultures (Wielkopolska, Silesia, Czech Republic) also occurred their linguistic full Indo-Europeanization and the emergence of centum dialects, mainly in Western Europe. It can not be ruled out that the Indo-European languages in the European branches of the haplogroup R1b already somehow stuck in the haploroup R1, paternal for R1a and R1b.
(Reflection. Archaeologists note a surprising „the phenomenon of the place” in the proglacial river Bachorza on the Kuyavia, especially with the towns: Dęby, Dobre and Smarglin. In this small area „extraordinary wealth of diverse cultural traditions” has been recorded, from the Proto-Neolithic age, through Neolithic, late Neolithic, Neolithic-Bronze interstadium to bronze in the Lusatian culture inclusive. A dozen well-known in Poland archaeological cultures or groups from these ages have been identified here, many of them betrays the other regions on the its earliness, suggesting, supposedly the local population on the Bachorza was the initiator or at least very active in taking from closer or further other people’s cultural initiatives – and to the extent that sometimes it is difficult to indicate their proper initiator and creator; see: Czebreszuk, Makarowicz 1990; Czebreszuk, Ignaszak, Łoś 1997)
………………………………………
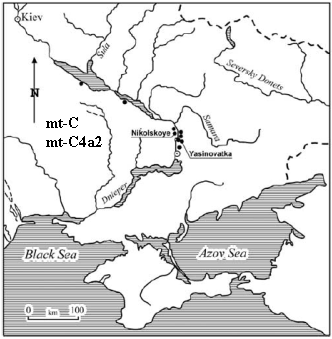
Haplogrupa mt-C (2 C i 1 C4a2) w neolitycznych grobach
w Nikolskoje i Jasinovovka nad Dnieprem
wg. A.G. Nikitin 2012,
http://eurogenes.blogspot.com/2012/06/ancient-mtdna-from-dnieper-donets.html
Matczyne grupy mt-C i jej podgrupy towarzyszyły ojcowskiej grupie R1a1-M459
w migracji ku Europie.
Zidentyfikowano je w archeologicznych szczątkach nad Dnieprem
i na na Jeleniej Wyspie na jeziorze Onega.
II.
Praindoeuropejczycy i ich wspólna ojczyzna
* Południowo-wschodnioazjatyckie pochodzenie R1a i języka pre-praindoeuropejskiego?
* Społeczność genetycznego rodu R1a-M417 w kręgu archeologicznej kultury rudoostrowsko-janisławickiej oraz w kulturze amfor kulistych na Kujawach i ziemiach polskich.
* Migracje Indoeuropejczyków w kolejnych kulturach horyzontu sznurowego i dialektach PIE
II-a.
Pre-praindoeuropejski ród (genetyczna haplogrupa)
R1a-M420 i jego migracja z południowo-wschodniej Azji ku Polsce
Haplogrupa to grupa ludzi mająca wspólnego przodka, u którego powstała określona mutacja ( zwana SNP, single nucledotide polimorphism). Mutacje w Y-DNA dziedziczone są w liniach prostych przez wszystkich męskich potomków określonego tą mutacją przodka. Według tymczasowych obliczeń tabeli, około 34.000 lat temu w genealogicznym drzewie człowieka współczesnego w ojcowskim chromosomie Y wyłoniła się mutacja-polimorfizm SNP, oznaczona symbolem M207, która – dziedziczona przez wszystkich męskich potomków – utworzyła ludzką grupę rodową, czyli haplogrupę, oznaczoną symbolem R. Język tej grupy można umownie nazwać dialektem języka nostratyckiego. Kolejny polimorfizm typu SNP, do którego mogło dojść około 33.000 lat temu także w Azji Południowo-Wschodniej dał początek rodu (haplogrupy) o symbolu R1-M173. Kolejna, jakby przełomowa mutacja, powstała około 29.000 lat temu; oznaczona R1a-M420 (jak na poniższym drzewie).
{kind=link}
- Sundajski epizod haplogrupy (rodu) R, R1a i R1a-M420
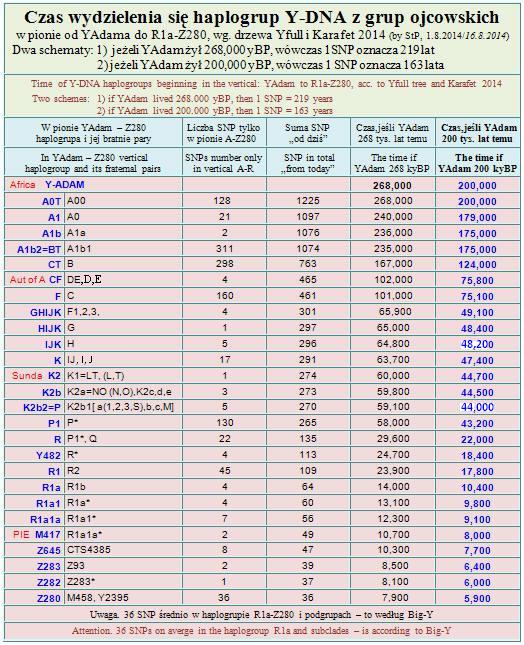
Kiedy i gdzie żył pierwszy ojciec wspólny dla całej gałęzi R1a? Według eksperymentalnego obliczenia czasu haplogrup współczesnego człowieka (tabela z 01.05.2014) gałąź R1a wyłoniła się z ojcowskiej R1 (i wcześniejszej R) orientacyjnie około 10,400 lat przed Chr.
Natomiast testowanie archeologicznego Y-DNA Malta-1 (MA-1) z południowej Syberii datuje R* (bez nowszych mutacji) na czas 24.000 lat temu.
Gdzie to nastąpiło? Po opublikowaniu 4 czerwca 2014 r. pracy Karafet et al. 2014 „Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia” http://dienekes.blogspot.com/2014/06/refined-structure-in-haplogroup-k-m526.html Już więc wiemy, że na drodze migracji naszych przodków z Afryki do Europy i Polski zaistniał ważny „epizod sundajski”.
{kind=link}
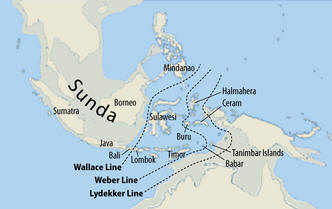
Otóż w czasie Wielkiego Zlodowacenia (LGM), gdy poziom oceanu był niższy nawet do 140 m, na terenie wielkiego wówczas lądu, zwanego Sunda (Sundaland), który był południowo-wschodnim przedłużeniem kontynentu azjatyckiego ku Australii, powstała (wyłoniła się) około 47.400 lat temu haplogrupa K-M526. Intensywnie tam, na Sundalandzie, rozwijająca się haplogrupa K-M526 wyłoniła z siebie wiele podgrup. Między nimi powstała haplogrupa N (dziś głównie w północnej Eurazji), O (dziś głównie w Azji wschodniej, Chiny), P i P1, Q (dziś Azja wschodnia i Ameryka), a w końcu R Grupy te wczesną migracją na północ i zachód Azji, jeszcze przed katastrofą Sundalandu, uratowały swoją wielką przyszłość.
{kind=link}
{kind=link}

Około 27.700 lat temu w rodzie R powstały podgrupy: euroazjatycka R1 i południowoazjatycka R2; następnie w tej pierwszej około 24.400 lat temu (22.400 przed Chr) powstały kolejne dwie podgrupy: R1a-M420 i R1b-M343. Język wczesnych etapów rozwoju grup R1a i R1b.
Trwająca kilka tysięcy lat migracja rodu R1a-M420* z Azji Południowo-Wschodniej ku Europie w ostatnim odcinku prowadziła zapewne, jak świadczą genetyczne ślady, przed północne Płaskowyże Irańskie
- W Azerii na północnym Płaskowyżu Irańskim czasowy pobyt rodu R1a-M420*?
Gdy po okresie LGM około 12.000 lat przed Chr. zaczęły kurczyć się brzegi Morza Kaspijskiego, odsloniły się i zazieleniły nadmorskie równiny na wschodzie, w pobliskich jaskiniach górskich mogła pojawić się jakaś pojawić się jakaś pierwsza ludność paleolityczna, a później protoneolityczna, z hodowlą owiec i kóz i ceramicznymi naczyniami na mleko. Co to za ludzie? Intuicyjnie i na podstawie dalszych archeologicznych śladów myślimy o migrującym rodzie R1a-M420*.
Na południowym wybrzeżu Morza Kaspijskiego w grotach Hotu i Belt w Iranie, u stóp irańskich gór, pojawiła się jakaś wczesna, przedrolnicza ludność, zostawiając po sobie zabytki kamienne i ceramiczne, świadczące o kontaktach z przodującymi ośrodkami rodzącego się neolitu u zboczy gór Zagros i w rejonie Żyznego Półksiężyca. Wyobraźnia każe domyślać się rodu R1a-M420*

Groty Belt i Hotu nad Morzem Kaspijskim w Iranie
więcej czytaj u ksiedza Stanisława Pietrzaka na http://www.tropie.tarnow.opoka.org.pl/polacy1.htm#Praindoeuropejczycy